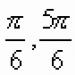Respiracijski koeficijent je omjer ugljičnog dioksida oslobođenog tijekom disanja i količine apsorbiranog kisika (CO2/O2). U slučaju klasičnog disanja, kada se ugljikohidrati CbH^O^ oksidiraju i kao konačni produkti nastaju samo CO2 i H2O, respiratorni koeficijent je jednak jedinici. Međutim, to nije uvijek slučaj, u nekim slučajevima se mijenja prema gore ili prema dolje, zbog čega se vjeruje da je pokazatelj produktivnosti disanja. Varijabilnost vrijednosti respiratornog koeficijenta ovisi o supstratu disanja (oksidirana tvar) i o produktima disanja (potpuna ili nepotpuna oksidacija).
Pri korištenju masti, koje su manje oksidirane od ugljikohidrata, umjesto ugljikohidrata tijekom disanja će se više kisika koristiti za njihovu oksidaciju - u tom slučaju će se respiratorni koeficijent smanjiti (na vrijednost od 0,6 - 0,7). To objašnjava veći sadržaj kalorija u mastima u usporedbi s ugljikohidratima.
Ako se tijekom disanja oksidiraju organske kiseline (tvari koje su jače oksidirane od ugljikohidrata), tada će se potrošiti manje kisika od oslobođenog ugljičnog dioksida, a respiratorni koeficijent će se povećati na vrijednost veću od jedan. Bit će najveći (jednak 4) tijekom disanja zbog oksalne kiseline, koja se oksidira prema jednadžbi
2 S2N2O4 + 02 4S02 + 2N20.
Gore je spomenuto da je uz potpunu oksidaciju supstrata (ugljikohidrata) u ugljični dioksid i vodu respiratorni koeficijent jednak jedan. Ali s nepotpunom oksidacijom i djelomičnim stvaranjem produkata poluživota, dio ugljika će ostati u biljci bez stvaranja ugljičnog dioksida; Apsorbirat će se više kisika, a respiratorni kvocijent će pasti na manje od jedinice.
Dakle, određivanjem respiratornog koeficijenta, može se dobiti predodžba o kvalitativnom smjeru disanja, supstratima i produktima ovog procesa.
Ovisnost disanja o čimbenicima okoliša.
Disanje i temperatura
Kao i drugi fiziološki procesi, intenzitet disanja ovisi o nizu čimbenika okoline, a jači je i
Najjasnije je izražena ovisnost o temperaturi. To je zbog činjenice da je od svih fizioloških procesa disanje "najkemijskiji", enzimski. Veza između aktivnosti enzima i razine temperature je neporeciva. Disanje se pridržava Van't Hoffovog pravila i ima temperaturni koeficijent (2ω 1,9 - 2,5.
Ovisnost respiracije o temperaturi izražena je jednovršnom (biološkom) krivuljom s tri kardinalne točke. Minimalna točka (zona) različita je za različite biljke. Kod hladno otpornih biljaka ono je određeno temperaturom smrzavanja biljnog tkiva, tako da se u nesmrzlim dijelovima četinjača disanje otkriva na temperaturama do -25 °C. Kod biljaka koje vole toplinu, minimalna točka je iznad nule i određena je temperaturom na kojoj biljke umiru. Optimalna točka (zona) za disanje nalazi se u rasponu od 25 do 35 °C, tj. nešto više od optimuma za fotosintezu. U biljkama s različitim stupnjem ljubavi prema toplini, njegov se položaj također donekle mijenja: nalazi se više u biljkama koje vole toplinu, a niže u onima otpornim na hladnoću. Maksimalna temperatura disanja je u rasponu od 45 do 53 °C.> Ova točka je određena smrću stanica i razaranjem citoplazme, jer stanica diše dok je živa. Dakle, temperaturna krivulja disanja slična je krivulji fotosinteze, ali je ne ponavlja. Razlika između njih je u tome što krivulja disanja pokriva širi temperaturni raspon od krivulje fotosinteze, a njen optimum je malo pomaknut prema višim temperaturama.
Oscilacije temperature snažno utječu na intenzitet disanja. Oštri prijelazi s visokog na nisko i natrag značajno povećavaju disanje, što je ustanovio * V. I. Palladin 1899. godine.
Pri fluktuacijama temperature dolazi do ne samo kvantitativnih, već i kvalitativnih promjena u disanju, tj. do promjena u putovima oksidacije organske tvari, ali one su za sada slabo proučene, pa ih ovdje ne prikazujemo.
Dah - jedan od najvažnijih metaboličkih procesa biljnog organizma. Energija koja se oslobađa tijekom disanja troši se kako na procese rasta tako i na održavanje aktivnog stanja biljnih organa koji su već završili rast. Međutim, važnost disanja nije ograničena samo na činjenicu da je to proces koji opskrbljuje energijom. Disanje, poput fotosinteze, složeni redoks proces, prolazeći kroz niz faza. U njegovim međufazama nastaju organski spojevi koji se potom koriste u raznim metaboličkim reakcijama. Intermedijarni spojevi uključuju organske kiseline i pentoze nastale različitim putevima respiratorne razgradnje. Tako, proces disanja je izvor mnogih metabolita.
Unatoč činjenici da je cjelokupni proces disanja suprotan fotosintezi, u nekim slučajevima oni se mogu nadopunjavati.
Oba procesa su dobavljači energetskih ekvivalenata (ATP, NADPH) i metabolita. Kao što se vidi iz sažete jednadžbe, voda nastaje i tijekom disanja. Tu vodu, pod ekstremnim uvjetima dehidracije, biljka može iskoristiti i zaštititi je od smrti. U nekim slučajevima, kada se energija disanja oslobađa kao toplina, disanje dovodi do nepotrebnog gubitka suhe tvari. U tom smislu, kada se razmatra proces disanja, mora se imati na umu da intenziviranje procesa disanja nije uvijek korisno za biljni organizam.
Pojedinačni stupnjevi disanja provode se u različitih dijelova biljne stanice. To je određeno raspodjelom enzima među pojedinim organelama s njihovim karakterističnim metaboličkim funkcijama. Proučavanje lokalizacije i topografije enzimskih sustava također je od velike važnosti za razumijevanje međudjelovanja pojedinih dijelova stanice, kao i mogućnosti međudjelovanja pojedinih metabolita.
Koncentriran u citoplazmi enzima koji kataliziraju proces glikolize i pentozofosfatnog puta. Postoje dokazi da su glikolitički enzimi također prisutni u mitohondrijskom matriksu. Enzimi Krebsovog ciklusa koncentrirani su uglavnom u matrici mitohondrija. Enzimi dišnog lanca utkani su određenim slijedom u unutarnju membranu mitohondrija. Otprilike 20-25% ukupnih proteina unutarnje membrane mitohondrija sastoji se od enzimskih proteina uključenih u prijenos protona i elektrona. Pretpostavlja se da enzimi nosači grupirani su tako da svaka skupina predstavlja samostalnu jedinicu – respiratorni ansambl. U mitohondrijima može biti nekoliko tisuća takvih skupina, koje su ravnomjerno raspoređene u membranama.
U unutarnja membrana mitohondrija Enzimi koji osiguravaju proces fosforilacije (ATP sintaza) također su lokalizirani. Tu je također koncentriran ATP transporter. Zahvaljujući tome, ATP nastao u mitohondrijima može ih napustiti i koristiti u drugim dijelovima stanice. Istodobno, isti transporter vrši prijenos ADP-a u unutarnji prostor mitohondrija. Pirogrožđana kiselina i neke organske kiseline Krebsovog ciklusa također prodiru kroz unutarnju membranu. Specifični transporter vrši prijenos fosfationa u mitohondrije. U isto vrijeme, unutarnja membrana je nepropusna za koenzime NAD i NADP i neke druge tvari.
Neki od koenzima nikotinamida obnavljaju se u citoplazmi tijekom glikolize. Da bi se izvršila njihova oksidacija, postoje posebni mehanizmi. Kod biljaka je NADH dehidrogenaza, pod čijim utjecajem NADH može ući u dišni lanac, lokalizirana na vanjskoj površini unutarnje membrane. U odsutnosti vanjske NADH dehidrogenaze, NADH se prenosi na unutarnju membranu pomoću mehanizma prijenosa. Suština ovog mehanizma je sljedeća. NADH stvoren u citoplazmi reagira s fosfodioksiacetonom, reducirajući ga u glicerofosfat. Glicerofosfat prodire kroz membranu i prenosi vodik do flavin dehidrogenaze i preko nje u dišni lanac. U tom slučaju, glicerofosfat se ponovno pretvara u fosfodioksiaceton, koji napušta mitohondrije u citoplazmi i ponovno se podvrgava redukciji NADH + H+. Sličan mehanizam za prijenos energetskih ekvivalenata kroz membrane također je pronađen u kloroplastima. Dakle, stanica istovremeno provodi i raspodjelu tvari po različitim odjeljcima i odnos među njima.
Važno je pitanje kako procesi koji se odvijaju u staničnoj jezgri opskrbljeni su energijom. Očigledno, ATP djelomično ulazi u jezgru iz citoplazme. Jezgra također sadrži vlastite dišne enzime. Tako su u nukleoplazmi pronađeni glikolitički enzimi. Postoje dokazi da enzimi respiratornog lanca slični enzimima mitohondrija djeluju u jezgri. Konačno, respiratorni enzimi također se nalaze u kloroplastima.
Supstrati za disanje biljaka
U radovima I.P. Borodin (1876) je pokazao da je intenzitet procesa disanja izravno proporcionalan sadržaju ugljikohidrata u biljnim tkivima. To je dalo razloga za pretpostavku da jest ugljikohidrati su glavna tvar koja se troši tijekom disanja (supstrat). U razjašnjenju ovog pitanja od velike je važnosti određivanje respiratornog koeficijenta. Respiracijski koeficijent (RK)- ovo je volumetrijski ili molarni omjer CO2 oslobođenog tijekom disanja prema apsorbiranom O2 tijekom istog vremenskog razdoblja, vrijednost DC ovisi o supstratu disanja. Ako se u procesu disanja koriste ugljikohidrati, tada se proces odvija prema jednadžbi
S6N1206 +602-> 6S02 + 6N20.
U ovom slučaju, DC je jednak jedan: 6S02/602 = 1.
Međutim, ako se više oksidiranih spojeva, na primjer, organskih kiselina, podvrgne raspadu tijekom disanja, apsorpcija kisika se smanjuje, DC postaje veći od jedinice. Dakle, ako se jabučna kiselina koristi kao supstrat za disanje, tada je DC = 1,33.
Određivanje respiratornih koeficijenata različitih biljnih tkiva pokazuje da je u normalnim uvjetima blizu jedinici. To daje razloga vjerovati da biljka prvenstveno koristi ugljikohidrate kao dišni materijal. Ako nedostaje ugljikohidrata, mogu se koristiti i drugi supstrati. To je posebno vidljivo kod klijanaca koje se razvijaju iz sjemenki koje sadrže masti ili bjelančevine kao rezervno hranjivo. U tom slučaju respiratorni koeficijent postaje manji od jedinice. Kada se masti koriste kao respiratorni materijal, one se razgrađuju na glicerol i masne kiseline. Masne kiseline mogu se pretvoriti u ugljikohidrate kroz ciklus glioksilata. Korištenju proteina kao supstrata za disanje prethodi njihova razgradnja na aminokiseline.
Supstrat, mješavina komadića treseta i posteljnog treseta, dobro je opskrbljen zrakom. Bez obzira na to koliko je supstrat mokar, korijen biljke ipak dobiva dovoljno kisika za disanje. Osim toga, treset se teško razgrađuje. Čak i uz jaku vlagu i visoku temperaturu, malo je vjerojatno da će istrunuti.[...]
Disanje je najnapredniji oblik oksidativnog procesa i najučinkovitiji način dobivanja energije. Glavna prednost disanja je u tome što se energija oksidirane tvari - supstrata na kojem mikroorganizam raste - najpotpunije koristi. Stoga se tijekom procesa disanja mnogo manje supstrata obrađuje za dobivanje određene količine energije nego, primjerice, tijekom fermentacije.[...]
Disanje se odnosi na proces povezan s razgradnjom ugljikohidrata, pri čemu se oslobađa energija koja osigurava metabolizam i transport u biljci. Budući da je kinetika metabolizma i transporta već opisana, troškovi supstrata za disanje mogu se izračunati iz poznatih ravnotežnih odnosa. Imajte na umu da se pri opisivanju disanja spajaju dvije faze pretvorbe kemijske energije: faza oksidacije supstrata, tijekom koje se stvaraju visokoenergetske ATP veze, i faza korištenja ATP energije. Osim toga, jednadžba ravnoteže disanja uzima u obzir trošak ugljikohidrata za osiguravanje energije za proces biosinteze i transporta organskih i anorganskih tvari. Tijekom disanja oslobađa se ugljikov dioksid koji se djelomično koristi u fotosintezi. Njegova dinamika opisana je na temelju ravnotežnih odnosa.[...]
Razlika u disanju obiju skupina fetusa, prema Helmeu i sur., vjerojatno je samo relativna (vidi odjeljak 1.3.4). Obje skupine imaju iste enzime i respiratorne supstrate. Razlog diferencijacije procesa disanja je, očito, kako u nejednakim citološkim promjenama tako iu nedovoljnoj aktivnosti enzima pojedinih reakcija [...]
Dakle, tijekom disanja, konačni akceptor vodika je kisik. Kod anaeroba, ili organski supstrati (fermentacija) ili anorganske tvari, kao što su nitrati ili sulfati ("anaerobno disanje") djeluju kao akceptori vodika. Dijagram pokazuje da se najjednostavniji i najprimitivniji prijenos elektrona provodi kod većine anaeroba zbog nepostojanja enzima u lancu transporta elektrona koji mogu prenijeti elektrone duž lanca do molekularnog kisika.[...]
Tijekom cijelog ljeta supstrat se održava toliko vlažnim da se iz šake uvijek bez puno truda može iscijediti nekoliko kapi tekućine. Visoka vlaga već će otežati disanje korijena, pa nakon svake jače kiše rub folije trebate nakratko spustiti i pustiti da višak vode iscuri.[...]
Već u trenutku vidljivih simptoma može se otkriti povećanje brzine disanja u listovima nekoliko sorti paprike (Capsicum sp.) zaraženih jakim sojem virusa graviranja, a visoka brzina disanja ostaje iu budućnosti. Drugačija je situacija s disanjem korijena oboljelih biljaka. Virus nije utjecao na brzinu disanja kod onih sorti kod kojih nije izazvao simptome venuća. Istodobno, kod inokulacije paprike Tabasco, koja je reagirala na infekciju virusom venuća, došlo je do smanjenja intenziteta disanja korijena 12-24 sata nakon povećanja propusnosti korijena (vidi str. 255). Pretpostavlja se da je smanjenje disanja u ovom slučaju uzrokovano istjecanjem supstrata i aktivatora enzima.[...]
Dakle, najjednostavniji proces aerobnog disanja predstavljen je u sljedećem obliku. Molekularni kisik koji se troši tijekom disanja koristi se uglavnom za vezanje vodika koji nastaje tijekom oksidacije supstrata. Vodik iz supstrata prelazi u kisik nizom intermedijarnih reakcija koje se odvijaju uzastopno uz sudjelovanje enzima i prijenosnika. Takozvani respiratorni koeficijent daje određenu predodžbu o prirodi procesa disanja. To se podrazumijeva kao omjer volumena oslobođenog ugljičnog dioksida i volumena apsorbiranog kisika tijekom disanja (C02:02).[...]
Tetrazolijeve soli također su se koristile kao supstrat za određivanje aktivnosti dehidrogenaze u tumorskim stanicama (Kraus, 1957.), za određivanje broja živih BCG bakterija u cjepivu (Eidus E. A., 1958.), za vitalno bojenje bakterija (Eidus E. A., 1959.). ), za detekciju mikroba otpornih na toplinu u mlijeku (Leali, 1958.), za prepoznavanje stanica kvasca s normalnim dišnim sustavom i s poremećenim disanjem (Ogur, 1957.) itd. Korištenjem dijagnostičkog medija koji sadrži tetrazolijeve soli, bakterije iz skupine Pseudomonas mogu se razlikovati (Selenka, 1958) i fitopatogene bakterije (Lovrekovich, Klement, 1960).[...]
V.I. Palladin je prvi razmatrao disanje kao niz enzimskih reakcija. U procesu oksidacije, oi je glavnu važnost pridavao procesu uklanjanja vodika iz supstrata.[...]
Ugljikohidrati su konačni produkt fotosinteze i supstrat za disanje i rast biljaka. Postoje podaci o zaštitnoj ulozi šećera u prilagodbi biljaka na nepovoljne uvjete okoliša (Kolupaev, Trunova, 1992). Svrha našeg istraživanja bila je ispitati sadržaj ugljikohidrata u biljkama ječma (Hordeum distichum L., selo Novichok) ovisno o stupnju mineralne ishrane i temperaturi. U pokusima smo koristili biljke stare 3-4 tjedna uzgojene u klima komori na vodenoj kulturi pri dva temperaturna režima (dan/noć) - niskom (13/8°C) i optimalnom (22/18°C) . Mineralni elementi svakodnevno su dodavani podlozi u eksponencijalno rastućim količinama kako bi se osigurala konstantna niska - 0,05 i visoka - 0,22 g/g-dnevna stopa rasta (Ingestad, Lund, 1986).[...]
Sada je pokazano da se u procesu disanja aktiviraju i vodik iz supstrata i kisik iz zraka.[...]
Vonrosa o tvarima koje se koriste u procesu disanja dugo su zaokupljali fiziologe. Još u djelima I. II. Borodin je pokazao da je intenzitet procesa disanja izravno proporcionalan sadržaju ugljikohidrata u biljnim tkivima. To je dalo razloga za pretpostavku da su ugljikohidrati glavna tvar koja se troši tijekom disanja. U razjašnjenju ovog pitanja od velike je važnosti određivanje respiratornog koeficijenta. Ako se u procesu disanja koriste ugljikohidrati, tada se proces odvija prema jednadžbi SeH 120b + 6O2 = 6CO2 + 6H2O, u tom slučaju respiratorni koeficijent je jednak jedinici - p = 1. Međutim, ako više oksidiranih spojeva, npr. organske kiseline, podvrgavaju se razgradnji tijekom procesa disanja, apsorpcija kisika se smanjuje, respiratorni koeficijent postaje veći od jedan. Kada se reducirani spojevi, poput masti ili proteina, oksidiraju tijekom disanja, potrebno je više kisika i respiratorni koeficijent postaje manji od jedinice.[...]
Pitanjem utjecaja svjetlosti na intenzitet disanja bavili su se mnogi fiziolozi. Rješenje ovog pitanja kompliciraju metodološke poteškoće. Na svjetlu je teško odvojiti proces fotosinteze od procesa disanja. Teško je razlikovati izravne i neizravne učinke svjetlosti. Dakle, fotosinteza se odvija na svjetlu, povećava se sadržaj ugljikohidrata - to utječe na proces disanja. Ipak, korištenje metode obilježenih atoma omogućilo je, iako ne u potpunosti, razlikovanje procesa fotosinteze od disanja. Trenutno se smatra da je utjecaj svjetlosti na proces disanja raznolik. Pod utjecajem svjetlosti, osobito kratkovalnih plavoljubičastih zraka, pojačava se intenzitet normalnog disanja u tami. Aktivacija disanja svjetlom dobro je dokazana na biljkama bez klorofila. Moguće je da swot aktivira enzime oksidaze. Svjetlost može posredno utjecati na disanje zelenih biljaka zbog odvijanja procesa fotosinteze. Zauzvrat, utjecaj fotosinteze na disanje može biti različit, pa čak i suprotan. Dakle, s jedne strane, tijekom procesa fotosinteze nastaju glavni supstrati disanja - ugljikohidrati. Zajedno s t[...]
Shema 1 (tablica 36) prikazuje transport elektrona tijekom disanja i različite vrste anaerobne proizvodnje energije. Vodik i elektroni uklanjaju se iz supstrata pomoću enzima piridin nukleotida (PN). Tok elektrona je usmjeren od sustava s nižim (negativnijim) potencijalom prema sustavu s višim (pozitivnijim) potencijalom, od - 0,8 - 0,4 V (potencijal supstrata) do +0,8 V (potencijal kisika).[ . ..]
Metoda za procjenu kinetike dodatne potrošnje supstrata od strane mikrobne populacije za izračunavanje heterotrofne aktivnosti u početku ne uzima u obzir mogući gubitak obilježenog ugljika zbog proizvodnje ugljičnog dioksida tijekom razdoblja inkubacije. Utvrđeno je da se, ovisno o vrsti supstrata, 8-60% primijenjenog obilježenog ugljika može izgubiti disanjem tijekom čak 3 sata inkubacije.[...]
Mehanizam uklanjanja iz otopine i naknadne disimilacije supstrata je vrlo složen i višefazan po prirodi, međusobno povezane i sekvencijalne biokemijske reakcije određene tipom prehrane i disanja bakterija.[...]
Ozljede biljnih organa i tkiva povećavaju intenzitet disanja. To može biti posljedica razaranja stanica, što povećava kontakt dišnih supstrata i enzima. Djelomično ranjavanje može uzrokovati ulazak stanica u meristematsku fazu rasta. Brzina disanja stanica koje se dijele uvijek je veća u usporedbi s onima koje su završile rast.[...]
Mnogi jednostavni fenoli utječu na energiju sustava i oksidaciju supstrata tijekom staničnog disanja.[...]
Relativna uloga ovih dišnih putova može varirati ovisno o vrsti biljke, starosti, razvojnoj fazi, kao i ovisno o uvjetima okoliša. Proces disanja biljaka odvija se u svim uvjetima u kojima je moguć život. Biljni organizam nema prilagodbe za regulaciju temperature, pa se proces disanja odvija na temperaturama od -50 do +50°C. Biljke također nemaju prilagodbe za održavanje ravnomjerne distribucije kisika u svim tkivima. Upravo je potreba za odvijanjem procesa disanja u različitim uvjetima dovela do razvoja u procesu evolucije različitih respiratornih metaboličkih putova i do još veće raznolikosti enzimskih sustava koji provode pojedine faze disanja. Važno je uočiti međusobnu povezanost svih metaboličkih procesa u biljnom organizmu. Promjena respiratornog metaboličkog puta dovodi do dubokih promjena u cjelokupnom metabolizmu biljnih organizama.[...]
Sadržaj vode. Lagani deficit vode rastućih tkiva povećava intenzitet disanja. To je zbog činjenice da nedostatak vode, pa čak i venuće lišća, pojačava razgradnju složenih ugljikohidrata (škroba) u jednostavnije (šećere). Povećanje sadržaja šećera u ovom glavnom supstratu disanja pospješuje sam proces. Istodobno, s nedostatkom vode, poremećena je veza oksidacije i glačanja fosfora. Disanje pod ovim uvjetima u biti je rasipanje suhe tvari. Duljim venućem biljka troši šećer i disanje se smanjuje. Drugačiji je obrazac tipičan za organe koji miruju. Povećanje sadržaja vode u sjemenu s 12 na 18% već povećava brzinu disanja za 4 puta. Daljnji porast udjela vode na 33% dovodi do povećanja brzine disanja za približno 100 puta. Kada se biljka ili tkivo premjesti iz vode u otopinu soli, ubrzava se disanje – to je takozvano disanje soli [...]
Nedostatak vode također mijenja osnovne fiziološke procese kao što su fotosinteza i disanje. Prije svega, kada se dehidrira, puči se zatvaraju, što naglo smanjuje protok ugljičnog dioksida u list i, kao posljedica toga, smanjuje se intenzitet fotosinteze. Međutim, smanjenje sadržaja vode smanjuje i intenzitet fotosinteze kod biljaka koje nemaju puči (mhp, lišajevi). Čini se da dehidracija, mijenjajući konformaciju enzima uključenih u proces fotosinteze, smanjuje njihovu aktivnost. To je zbog činjenice da se kao rezultat brze razgradnje škroba pod utjecajem sušenja povećava količina šećera - ovog glavnog supstrata disanja. Istodobno, pri nedostatku vode u stanicama, energija koja se oslobađa tijekom disanja ne akumulira se u ATP-u, već se uglavnom oslobađa u obliku topline. Zbog toga pojačano disanje, praćeno razgradnjom organskih tvari, može štetiti biljnom organizmu.[...]
Najčešće pojavu plijesni uzrokuju gljivice iz rodova Micor, Aspergillus, Dematium koje žive na raznim podlogama i vrlo su česte u prirodi. Plijesni su najosjetljivije sjemenke koje sadrže velike količine vode, oštećene su ili su pohranjene u uvjetima visoke vlažnosti. Šteta od plijesni leži u činjenici da gljive obavijaju sjeme micelijem, ometaju disanje i druge fiziološke procese koji se odvijaju u sjemenu tijekom skladištenja i često uzrokuju njihovu smrt. Ponekad pljesnivo sjeme niče, ali se sporo razvija i u pravilu je jako pogođeno raznim uzročnicima [...]
Denitrifikacija, kao mikrobiološki proces, samo je poseban oblik disanja u slučaju nedostatka kisika. Razne bakterije u biološkom postrojenju za pročišćavanje otpadnih voda, uglavnom proteolitičke bakterije, mogu osigurati redukciju dušika i nitrata u odsutnosti slobodnog kisika i u prisutnosti odgovarajućeg supstrata izvora vodika. Dakle, kemijski vezani kisik može se koristiti za metaboličke procese ovih bakterija. Sposobnost denitrifikacije bakterije stječu tijekom procesa prilagodbe. Izvor ugljikovodika mora se dozirati u minimalnom omjeru koji odgovara sadržaju nitrata.[...]
Budući da je oksidacija dio svakog aerobnog procesa, to znači da se organski supstrat nikada ne može pretvoriti 100% u organsku tvar biomase. Naravno, ako se uzme u obzir proizvodnja ugljičnog dioksida, neće biti gubitka ugljika. U procesima nakupljanja rezervnih tvari povećanje može doseći 0,95 g KPK/g KPK(B). Druga ekstremna situacija je da se cijeli supstrat troši na održavanje vitalne aktivnosti stanica (endogeno disanje), kao rezultat toga, povećanje biomase je nula ili čak negativno. Za određenu količinu supstrata, povećanje biomase ovisi o trajanju procesa.[...]
Uključivanje (Eyu u ekstrahirane mitohondrije dovodi do značajnog povećanja supstratom induciranog signala (u prosjeku za 80%) i obnavlja njegovu ovisnost o Fn, ali ne i o ADP. Uvođenje (Eyu također je dovelo do značajnog povećanja u disanje, ali ne i obnova fosforilacije, koja je povezana, očito, s poremećajem membranskih struktura tijekom ekstrakcije ubikinona.[...]
Disimilacija ugljikohidrata može se dogoditi na dva načina. U plodovima jabučica šećer se uglavnom troši na disanje duž EMP (Embden - Meyerhof - Parnas) putanje. U tom slučaju, zbog procesa fosforilacije, glukoza se razgrađuje u pirogrožđanu kiselinu (glikoliza). Osim toga, postoji mogućnost razgradnje ugljikohidrata kroz pentozni ciklus. U kojoj je mjeri ovaj ciklus uključen u transformaciju dišnih supstrata još se ne može reći. Trenutno se pretpostavlja da u određenim fazama razvoja jabuke ili drugog voća prevladava jedan ili drugi put. EMT put koji prevladava u jabučastom voću završava s pirogrožđanom kiselinom, koja ima ključnu ulogu u disanju. Od tog trenutka daljnje transformacije pirogrožđane kiseline ovise o okolini: u aerobnim - uz potrošnju kisika, u anaerobnim - kada kisik nije potreban.[...]
Na temelju prirode disimilacije razlikuju se aerobni i anaerobni organizmi. Aerobni (od grč. aeg - zrak) organizmi koriste slobodni kisik za disanje (oksidaciju). Većina živih organizama su aerobi. Naprotiv, anaerobi oksidiraju supstrate, poput šećera, u nedostatku kisika, stoga je za njih disanje fermentacija. Mnogi mikroorganizmi i helminti su anaerobi. Na primjer, dinitrifikacijske anaerobne bakterije oksidiraju organske spojeve pomoću nitrita, koji je anorgansko oksidacijsko sredstvo.[...]
Kao što je već navedeno, mnoge skupine bakterija (na primjer, fakultativni anaerobi) sposobne su i za aerobno i za anaerobno disanje, ali krajnji produkti tih dviju reakcija su različiti i količina energije koja se oslobađa tijekom anaerobnog disanja je mnogo manja. Na sl. Slika 2.7 prikazuje rezultate zanimljive studije u kojoj je ista vrsta bakterije, Aegobacler, uzgajana u anaerobnim i aerobnim uvjetima koristeći glukozu kao izvor ugljika. U prisutnosti kisika gotovo sva glukoza pretvorena je u bakterijsku biomasu i CO2; u nedostatku kisika, razgradnja je bila nepotpuna, mnogo manji dio glukoze pretvoren je u stanične tvari koje sadrže ugljik, a niz organskih spojeva otpušten je u vanjski okoliš. Za njihovu oksidaciju bile bi potrebne druge specijalizirane vrste bakterija. Kada je stopa unosa organskog detritusa u tlo i sedimente velika, bakterije, gljive, protozoe i drugi organizmi stvaraju anaerobne uvjete koristeći kisik brže nego što on difundira u supstrat. Istodobno, razgradnja organske tvari ne prestaje - ona se nastavlja, iako često sporijim tempom, ako okoliš sadrži mikroorganizme s prilično širokim rasponom anaerobnih vrsta metabolizma.[...]
Vrijednost DC ovisi o drugim razlozima. U nekim tkivima, zbog otežanog pristupa kisika, uz aerobno disanje javlja se i anaerobno disanje koje nije popraćeno apsorpcijom kisika, što dovodi do porasta vrijednosti DC. Vrijednost koeficijenta također je određena potpunošću oksidacije dišnog supstrata. Ako se uz finalne produkte u tkivima nakupljaju i manje oksidirani spojevi (organske kiseline), tada DC[...]
Dinamika oslobađanja ugljičnog dioksida (S?SO2), apsorpcija kisika ([...]
Čim voda počne teći u sjemenke, njihovo disanje se naglo povećava, a istovremeno se aktiviraju različiti enzimi koji nastaju tijekom razdoblja zrenja. Pod njihovim utjecajem rezervne hranjive tvari se hidroliziraju, pretvarajući se u pokretni, lako probavljivi oblik. Masti i škrob se pretvaraju u organske kiseline i šećere, proteini u aminokiseline. Prelazeći u embrij iz skladišnih organa, hranjive tvari postaju supstrat za procese sinteze koji u njemu započinju, prvenstveno novih nukleinskih kiselina i enzimskih proteina neophodnih za početak rasta.
Metabolička priroda druge faze konzumacije također je potvrđena izostankom, nakon prve faze apsorpcije tvari, daljnjeg nakupljanja C14 u kulturi kada je disanje bakterija inhibirano uvođenjem kalij cijanida u supstrat.[.. .]
Pretjerano razvijena vegetacija ometa pravilan rad ribnjaka i pridonosi pogoršanju hidrokemijskih i plinskih prilika, osobito noću, kada kisik troše svi vodeni organizmi za disanje i stvara se njegov nedostatak. Pri raspadanju umiruće vegetacije oslobađaju se toksični produkti raspadanja (amonijak, sumporovodik i dr.), a njeni ostaci su supstrat za očuvanje i razmnožavanje saprofitskih i patogenih gljiva i bakterija.[...]
Za normalan tijek sinteze proteina u tijelu biljke potrebni su sljedeći uvjeti: 1) opskrba dušikom; 2) opskrba ugljikohidratima (ugljikohidrati su neophodni i kao materijal za izgradnju ugljičnog kostura aminokiselina i kao supstrat za disanje); 3) visok intenzitet i konjugacija procesa disanja i fosforilacije. U svim fazama transformacije dušičnih tvari (redukcija nitrata, stvaranje amida, aktivacija aminokiselina tijekom sinteze proteina itd.), potrebna je energija sadržana u visokoenergetskim fosfornim vezama (ATP); 4) prisutnost nukleinskih kiselina: DNA je neophodna kao tvar u kojoj su šifrirane informacije o slijedu aminokiselina u sintetiziranoj proteinskoj molekuli; mRNA - kao sredstvo koje osigurava prijenos informacija od DNA do ribosoma; t-RNA - kapa koja osigurava prijenos aminokiselina u ribosome; 5) ribosomi, strukturne jedinice u kojima se odvija sinteza proteina; 6) enzimski proteini, katalizatori za sintezu proteina (aminoacil-t-RNA spptetaze); 7) niz mineralnih elemenata (ioni Mg2+, Ca2+).[...]
U delti Volge, K.V Gorbunov (1955) identificirao je 4 razdoblja u razvoju obraštanja: embrionalni, brzi rast, umjerena aktivnost i smanjenje biomase. Broj vrsta i biomasa obraštaja dosegli su maksimum 10. dana. Do tog vremena smanjio se intenzitet fotosinteze i disanja, više od polovice biomase obraštanja činile su protozoe, rotiferi i bakterije. Prema drugim autorima (Cattaneo, 1975.), bilo je potrebno oko 4 tjedna da se obraštaj stabilizira i postigne njegova sličnost sa zajednicama na prirodnim supstratima.[...]
Svi fiziološki procesi u biljci odvijaju se normalno samo uz optimalnu opskrbljenost vodom. Voda nije samo otapalo, već i aktivna strukturna komponenta stanice. Uključen je u biološke transformacije, primjerice, olakšava interakciju među molekulama, služi kao supstrat za fotosintezu, sudjeluje u disanju i brojnim hidrolitičkim i sintetskim procesima.[...]
Među ostalim klasama kralješnjaka, ribe, osobito slatkovodne, možda češće nego druge, suočene su s nepovoljnim režimom kisika, osobito s njegovim sezonskim i dnevnim kolebanjima u vodenim tijelima, s akutnim nedostatkom zimi. Stoga, u pogledu prirode energetskog metabolizma i omjera specifične težine glikolize i disanja, ribe zauzimaju srednji položaj između fakultativnih anaeroba (beskralješnjaka) i tipičnih aeroba (viših kralježnjaka). Posebne studije su pokazale da ribe karakterizira smanjena razina oksidativnih procesa i smanjena aktivnost citokromskog sustava u usporedbi s toplokrvnim životinjama. Prema tim autorima, oksidacijski sustavi riba su "primitivniji" od onih viših kralješnjaka. Na primjer, aktivnost citokrom oksidaze kod pojedinih vrsta riba koštunjača je visoka, a sadržaj citokroma b je nizak. Osim toga, citokrom b nije jedini supstrat za riblju citokrom oksidazu. Posljedično, najvažniji respiratorni enzim, koji dovršava faze disanja, nema strogu specifičnost. Sustav oksidativne fosforilacije riba funkcionira manje učinkovito od sustava drugih skupina kralješnjaka.[...]
Pri analizi odnosa između veličine i metabolizma kod biljaka često nije lako odlučiti što se, strogo govoreći, smatra “jedinkom”. Dakle, veliko stablo se može smatrati jednom jedinkom, ali kada se proučava odnos između veličine i površine, lišće se može smatrati "funkcionalnim jedinkama" (sjetite se koncepta "indeksa površine lista"). Proučavajući različite vrste velikih morskih višestaničnih algi, otkrivamo da vrste s tankim ili uskim "granama" (tj. visokim omjerom površina/volumen) imaju višu razinu proizvodnje hrane po gramu biomase, intenzivnije disanje i apsorpciju radioaktivnog fosfora iz vode nego vrste s debelim "granama" (E. Odum, Kuentzler, Blunt, 1958). U ovom slučaju, "funkcionalne jedinke" su "grane" ili čak pojedinačne stanice, a ne cijela biljka, koja može biti formirana od mnogih "grana" pričvršćenih za podlogu jednim rizoidom.
Kod biljaka postoje dva puta oksidacije dišnog supstrata: glikoliza i pentozofosfatni put.
Glikoliza je anaerobni proces koji se odvija u citoplazmi. S biološke perspektive, glikoliza je vrlo primitivan proces koji je nastao prije pojave kisika u Zemljinoj atmosferi i formiranja staničnih organela.
U složenom lancu glikolitičke razgradnje ugljikohidrata mogu se razlikovati dvije karike (9 reakcija):
U prvoj vezi troši se ATP energija; u drugom se spojevi sa šest ugljika (fruktoza-1,6 difosfat) razbijaju uz stvaranje trioza; u trećem se energija skladišti (oslobađa). Hidrolizi nije podvrgnuta slobodna glikolitička molekula, već ona koju aktivira ATP. Ta se aktivacija naziva fosforilacija.
Kao rezultat fosforilacije nastaje glukoza-6-fosfat. Daljnja aktivacija heksoze postiže se pretvaranjem glukoza-6 fosfata u fruktoza 6 fosfat. U sljedećoj fazi, fruktoza-6 fosfatu se dodaje još jedan ostatak fosforne kiseline. Donator fosforne kiseline i energije potrebne za nastanak estera je molekula ATP. Reakcije prijenosa katalizira enzim fosfoheksokenaza. Rezultat ove reakcije je stvaranje fruktoza-1,6-bisfosfata.
U drugoj karici: rezultirajuća molekula fruktoza-1,6-difosfata razbija se na 3-fosfogliceraldehid i *. Reakciju rupture katalizira enzim aldolaza.
Samo fosfogliceraldehid dalje sudjeluje u procesima glikolitičke razgradnje. Fosfodioksiaceton se potpuno pretvara u fosfogliceraldehid. Fosfogliceraldehid se oksidira u 1,3-difosfoglicerinsku kiselinu.
U trećoj karici: nastala 1,3-difosfoglicerinska kiselina ulazi u enzimsku reakciju s ADP. Kao rezultat toga, jedna od njegovih fosfornih skupina se prenosi na ADP da bi se formirao ATP i 3-fosfoglicerinska kiselina.
Stvaranje ATP-a u citoplazmi tijekom enzimskih reakcija naziva se fosforilacija supstrata. 3FGK se pomoću enzima * pretvara u 2FGK. 2PHA se pretvara u 2-fosfoenolpirugrožđanu kiselinu pomoću enzima enolaze.
Uklanjanjem fosfornog ostatka iz PPVK nastaje enolPVC koji zbog svoje nestabilnosti spontano prelazi u ketokiselinu PPV.
Stvaranje PVC-a prolazi kroz daljnju razgradnju, i anaerobnu i aerobnu, u ciklusu di- i trikarboksilne kiseline. Anaerobna probava, tj. bez sudjelovanja O 2 PVC može nastati po tipu alkoholnog vrenja ili po tipu mliječno-kiselog vrenja. Tijekom alkoholnog vrenja nastaju etilni alkohol i CO 2 . Za mesnate, sočne plodove alkoholna fermentacija je normalan fiziološki proces. Za cijelu biljku ili za sustav konja, dugotrajna izloženost uvjetima nedovoljne prozračnosti, alkoholnog vrenja ima štetan učinak, što dovodi do smrti.
Zašto? Budući da je fermentacija popraćena oslobađanjem male količine energije, koja nije dovoljna za dugotrajno održavanje života, a nakupljanje alkohola dovodi do trovanja tijela. Anaerobno disanje tipa fermentacije događa se u uvjetima poplave.
U aerobnim uvjetima PVK u mitohondrijima potpuno se oksidira do CO 2 i H 2 O. Ta se oksidacija, kako je utvrdio engleski biokemičar Krebs, odvija u uzastopnim koracima uz nastajanje di- i trikarboksilnih kiselina. Krebsov ciklus može se podijeliti u tri dijela.
U prvom dijelu PVA se oksidira u octenu kiselinu uz stvaranje acetil CoA i oslobađanje CO2.
Drugi dio ciklusa započinje reakcijom između PCA i Acetyl CoA, što dovodi do sinteze limunske kiseline. Limunska kiselina se potom pretvara nizom intermedijarnih spojeva (izocitrična kiselina) u oksalno-jantarnu kiselinu. Oksalno-jantarna kiselina prolazi kroz dekarboksilaciju, što rezultira otpuštanjem CO 2 i stvaranjem X-ketoglutarne kiseline. X-ketoglutarna kiselina ponovno se dekarboksilira – oslobađa se CO 2 i nastaje jantarna kiselina. U ovom dijelu ciklusa octena kiselina se potpuno oksidira (oslobađanjem CO2) i time je završena oksidacija PVC-a.
Treći dio ciklusa predstavlja međupretvorbu dibazičnih kiselina s 4 ugljikova atoma - jantarna → fumarna → jabučna → i završava regeneracijom PIKE.
ATP se ne sintetizira izravno u Krebsovom ciklusu, isključujući supstratnu fosforilaciju X-ketoglutarne kiseline, ali se pet molekula reduciranih nukleotida pojavljuje u ciklusu:
1. tijekom oksidativne dekarboksilacije PVC-a;
2. tijekom dehidrogenacije izocitrične kiseline;
3. tijekom oksidacije ketoglutarne kiseline;
4. tijekom oksidacije jantarne kiseline;
5. tijekom oksidacije jabučne kiseline.
Svaki par vodikovih atoma (H +, e -) nakon eliminacije prelazi iz supstrata u kisik kroz niz nosača lokaliziranih u unutarnjoj membrani mitohondrija. Prijenos elektrona kroz ETC također je povezan sa sintezom ATP-a. Proces stvaranja ATP-a povezan s prijenosom elektrona kroz ETC mitohondrija naziva se oksidativna fosforilacija. Na kraju lanca, elektrone hvata kisik i spajaju se s protonima (ion zraka) u molekulu vode.
Koliki je prinos energije iz oksidacije glukoze? Tijekom procesa disanja, glikoliza (fosforilacija supstrata: 8 ATP molekula) i Krebsov ciklus (oksidativna fosforilacija proizvodi 30 ATP molekula) proizvode 38 ATP molekula. Učinkovitost korištenja energije kroz glikolizu i Krebsov ciklus je učinkovitost = 1596/2721*100% = 58,6%.
U biljnim stanicama, uz glikolizu i Krebsov ciklus, postoji još jedan put oksidacije ugljikohidrata - pentozofosfat. Oksidacija glukoze u ovom ciklusu povezana je s eliminacijom prvog (aldehidnog) ugljikovog atoma u obliku CO2. Početni proizvod u pentozofosfatnom ciklusu je glukoza-6-fosfat, koji se dalje oksidira u 6-fosfoglukonsku kiselinu.
U pentozofosfatnom ciklusu, ATP se koristi za stvaranje početnog produkta: za fosforilaciju glukoza-6-fosfata. Sve reakcije pentozofosfatnog puta odvijaju se u topivom dijelu stanične citoplazme, kao iu protoplastidima i kloroplastima. ATP se ne proizvodi niti u jednoj reakciji ovog ciklusa, ali ovaj ciklus je dobavljač vodika za ETC disanje. NADH služi kao donor vodika za ETC disanje. Energetski učinak PPP-a je 36 molekula ATP-a. Glavna svrha PFP-a je sudjelovanje ne toliko u energetskom, već u plastičnom metabolizmu. Pentozofosfatni put od velike je važnosti kao izvor stvaranja ugljikohidrata s različitim brojem ugljikovih atoma u lancu - od C3 do C7. PPP služi kao glavni ekstrakloroplastni i ekstramitohondrijski izvor NADPH, koji je neophodan za sintezu masnih kiselina.
Biološka uloga pentoza nužna za sintezu nukleotida, tj. za sintezu riboze i deoksiriboze. Pomak prema pentozofosfatnom putu događa se kada stanica zahtijeva velike količine šećera s pet ugljika i kada se NADPH koristi kao izvor energije za sintezu umjesto NADH.
Biljke primarno koriste ugljikohidrate kao dišni materijal. pri čemu respiratorni kvocijent(molarni omjer CO 2 oslobođenog tijekom disanja i O 2 apsorbiranog tijekom istog vremenskog razdoblja) jednak je jedan ().
Ako nedostaje ugljikohidrata, mogu se koristiti i drugi supstrati. To je posebno vidljivo kod klijanaca koji se razvijaju iz sjemenki koje sadrže bjelančevine ili masti kao rezervno hranjivo. Budući da su ti spojevi reduciraniji od šećera, njihova oksidacija zahtijeva više kisika i respiratorni koeficijent postaje manji od jedinice. U tom slučaju, masti se najprije razgrađuju na glicerol i masne kiseline, koje se kroz ciklus glioksilata mogu pretvoriti u ugljikohidrate. Korištenju proteina kao supstrata za disanje prethodi njihova razgradnja na aminokiseline.
Respiratorni putevi
Postoje dva glavna načina pretvaranja dišnog supstrata ili oksidacije ugljikohidrata:
Dihotomno: glikoliza + Krebsov ciklus;
Apotomski: pentozofosfatni put
Relativna uloga ovih respiratornih putova može varirati ovisno o vrsti biljke, starosti, fazi razvoja, a također i ovisno o uvjetima okoline (na primjer, disanje se događa u temperaturnom rasponu od -50...+50 0 C).
Razmotrimo oba načina:
glikoliza – proces anaerobne razgradnje glukoze, koji se javlja uz oslobađanje energije, čiji je krajnji produkt pirogrožđana kiselina (PVA).
Reakcije glikolize odvijaju se u topljivom dijelu citoplazme (citosolu), gdje se glikolitički enzimi organiziraju u multienzimske komplekse uz sudjelovanje aktinskih filamenata citoskeleta, te u kloroplastima.
Lanac reakcija koji čini put glikolize može se podijeliti u 3 faze:

Dakle, ukupna jednadžba za glikolizu je sljedeća:
Funkcije glikolize u stanici:

Krebsov ciklus. U anaerobnim uvjetima PVC prolazi različite vrste fermentacije. U prisutnosti dovoljnog kisika, piruvat se potpuno oksidira u ugljični dioksid i vodu u Krebsovom ciklusu. Svi sudionici u ovom procesu lokalizirani su u matrici ili u unutarnjoj membrani mitohondrija.
Prva faza oksidativnog cijepanja PVK je proces stvaranja aktivnog acetila tijekom oksidativne dekarboksilacije uz sudjelovanje multienzimskog kompleksa piruvat dehidrogenaze (3 enzima i 5 koenzima).
Kao rezultat, nastaje acetil koenzim A:
(acetil - Co A)

Daljnja oksidacija acetil-Co A odvija se tijekom cikličkog procesa (Krebsov ciklus) koji počinje interakcijom s oksalooctenom kiselinom (OA):
Kao rezultat jedne revolucije ciklusa:
Sažetak aerobne oksidacije:
Sažetak glikolize i Krebsovog ciklusa:
Glioksilatni ciklus (nema ga u životinjskim stanicama). Može se smatrati modifikacijom Krebsovog ciklusa. Aktivno djeluje u klijavim sjemenkama uljarica, gdje se skladišne masti pretvaraju u šećere (glukoneogeneza). Lokaliziran ne u mitohondrijima, već u specijaliziranim mikrotijelima - glioksisomi.
Za razliku od Krebsovog ciklusa, svaki promet uključuje ne jednu, već dvije molekule acetil-CoA A (nastale tijekom β-oksidacije masnih kiselina) i koristi se za sintezu jantarne kiseline, koja napušta glioksisome, pretvara se u ACA i uključuje u glukoneogenezi (reverzna glikoliza).

Put oksidacije pentoza fosfata (PPP, pentozni shunt). Oksidacija glukoze na tom putu povezana je s uklanjanjem prvog atoma ugljika iz glukoze u obliku CO2.
Taj se proces odvija u topivom dijelu stanične citoplazme i u kloroplastima. PPP respiracija posebno je aktivna u onim biljnim stanicama i tkivima u kojima su intenzivni sintetski procesi, poput sinteze lipidnih komponenti membrana, nukleinskih kiselina, staničnih stijenki i fenolnih spojeva.
Postoje 2 faze PFP-a:
oksidacija glukoze u ribuloza-5-fosfat uz stvaranje 2NADPH*H +;
rekombinacija šećera za regeneraciju izvornog supstrata uz sudjelovanje enzima transketolaza, transaldolaza i izomeraza.
Ukupna PFP jednadžba izgleda ovako:
6C 6 H 12 O 6 = 5 C 6 H 12 O 6 + 6CO 2 + 12NADPH*H +
Glavno značenje PPP-a nije energetsko, već plastično:
1 - NADPH*H +, koji se ne stvara u mitohondrijima, već u citoplazmi, koristi se uglavnom u raznim sintetskim procesima (jer, za razliku od NAD +, koji je u stanicama prisutan uglavnom u stabilnijem oksidiranom obliku, NADPH*H + je u reduciranom obliku).
2 – Tijekom PPP sintetiziraju se pentoze koje su dio nukleinskih kiselina i nukleotida (ATP, GTP, UTP, NAD, FAD, koenzim A su također nukleotidi, koji uključuju ribozu).
3 – Ugljikohidrati C 3 – C 7 potrebni su za sintezu raznih tvari. Na primjer, šikiminska kiselina se sintetizira iz eritroze, prekursora mnogih aromatskih spojeva, kao što su aminokiseline, vitamini, tanini i tvari za rast, lignin stanične stijenke itd.
4 – PFP komponente sudjeluju u tamnoj fiksaciji CO 2. Samo 2 od 15 reakcija Calvinovog ciklusa su specifične za fotosintezu, ostale su uključene u PPP.
5 – Trioze C 3 iz PPP mogu se pretvoriti u 3-PGA i sudjelovati u glikolizi.

Normalna aktivnost PPP u općoj respiratornoj izmjeni je 10-40%. Aktivnost se povećava u nepovoljnim uvjetima: suša, nedostatak kalija, infekcija, zasjenjenje, slanost, starenje.
Respiratorni transportni lanac elektrona i oksidativna fosforilacija.
Krebsov ciklus, glioksilat i PPP funkcioniraju samo u uvjetima dovoljne količine O2. U isto vrijeme, O2 nije izravno uključen u reakcije ovih ciklusa. Neophodan je za završnu fazu respiratornog procesa povezanog s oksidacijom akumuliranih reduciranih koenzima NADH*H+ i FADH 2 u respiratornom transportnom lancu elektrona (ETC) mitohondrija. Prijenos elektrona kroz ETC također je povezan sa sintezom ATP-a.
Respiratorni ETC, lokaliziran u unutarnjoj membrani mitohondrija, sastoji se od četiri multienzimska kompleksa, koji uključuju (kako redoks potencijal raste):
FMNželjezo sumpor proteiniFeS –ubikinonP –citokromi (b 556 ,b 560 , 1 c,c,a,a 3 ) - OKO 2 . U ovom slučaju, elektroni iz NADH*H + idu u FMN, a iz FADH 2 - izravno u ubikinon.
Iz mitohondrijskog matriksa, tijekom transporta svakog para elektrona od NADH*H + do O 2 u tri dijela ETC-a kroz membranu prema van (u međumembranski prostor), prenese se najmanje šest H + protona, tijekom oksidacija FADH 2 - samo dva takva odjeljka.
Kao rezultat, na membrani se stvara elektrokemijski potencijal H + iona, uključujući kemijski ili osmotski gradijent (ΔpH) i električni gradijent Prema Mitchellovoj kemiosmotskoj teoriji, ovaj elektrokemijski transmembranski potencijal H + iona je izvor energije. za sintezu ATP-a zbog transporta protona kroz protonski kanal membrane ATP – osn.
A  Kompleks TFase sastoji se od faktora spajanja F 1 (protein od 9 podjedinica) i faktora F o koji međusobno povezuje membranu i služi kao kanal za transport H + iona.
Kompleks TFase sastoji se od faktora spajanja F 1 (protein od 9 podjedinica) i faktora F o koji međusobno povezuje membranu i služi kao kanal za transport H + iona.
Proces fosforilacije ADP-a sa stvaranjem ATP-a, zajedno s prijenosom elektrona tijekom mitohondrijskog ETC-a, naziva se oksidativne fosforilacije .
Tijekom oksidacije NADH*H +, koeficijent fosforilacije = 3, FADH 2 = 2, odnosno ima dovoljno energije za sintezu 3, odnosno 2 molekule ATP.
C 6 H 12 O 6 = 6CO 2 + 4 ATP + 10 NADH*H + + 2 FADH 2.
U ovom slučaju, 10 NADH*H + = 30 ATP, a 2 FADH 2 = 4 ATP. Kao rezultat imamo 4 +30+4 =38 ATP = 380 kcal/mol = 1591 kJ/mol energije.