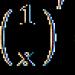пар іншими. Зазначені зміни справді супроводжують кожен цикл реплікації ДНК, проте їх частота значно менша, ніж мала б бути. Це тим, що більшість змін такого роду усувається завдяки дії механізму репарації (молекулярного відновлення) вихідної нуклеотидної послідовності ДНК.
Механізм репарації ґрунтується на наявності в молекулі ДНК двох комплементарних ланцюгів. Спотворення послідовності нуклеотидів в одній з них виявляється специфічними ферментами. Потім відповідна ділянка видаляється та заміщається новим, синтезованим на другому комплементарному ланцюзі ДНК. Таку репарацію називають ексцизійною, тобто. з "вирізанням" (рис. 3.15). Вона здійснюється до чергового циклу реплікації, тому її називають також
дореплікативною.
Мал. 3.14. Схема процесу корекції при синтезі ДНК: I - включення в ланцюг ДНК нуклеотиду із зміненою (таутомерною) формою цитоеїну, який «незаконно» спарується з аденіном; II - швидкий перехід
цитозину у звичайну форму порушує його спарювання з аденіном; неспарений 3"- ВІН-кінець синтезованого ланцюга перешкоджає подальшому її подовженню під дією ДНК-полімерази; III - ДНК-полімераза видаляє незаконний нуклеотид, в результаті чого знову з'являється спарений з матрицею 3" - ВІН-кінець; IV - ДНК-полімераза продовжує нарощування ланцюга на 3"-ВІН-кінці.
Відновлення вихідної структури ДНК потребує участі низки ферментів. Важливим моментом запуску механізму репарації є виявлення помилки в структурі ДНК. Нерідко такі помилки виникають у новоствореному синтезованому ланцюгу в процесі реплікації. Ферменти репарації повинні виявити саме цей ланцюг. У багатьох видів живих організмів знову синтезований ланцюг ДНК відрізняється від материнським ступенем метилювання її азотистих основ, що відстає від синтезу. Репарації при цьому неметильований ланцюг. Об'єктом впізнавання ферментами репарації можуть також служити розриви ланцюга ДНК. У вищих організмів, де синтез ДНК відбувається не безперервно, а окремими репліконами, знову синтезований ланцюг ДНК має розриви, що уможливлює її впізнавання.
Відновлення структури ДНК при втраті пуринових основ одного з її ланцюгів передбачає виявлення дефекту за допомогою ферменту ендонуклеази, що розриває фосфоефірний зв'язок у місці пошкодження ланцюга. Потім змінена ділянка з кількома нуклеотидами, що примикають до неї, видаляється ферментом екзонуклеазою, а на його місці відповідно до порядку підстав комплементарного ланцюга утворюється правильна нуклеотидна послідовність (рис. 3.15).

Мал. 3.15. Схема ексцизійної, дореплікативної репарації ДНК При зміні однієї з основ у ланцюзі ДНК у відновленні вихідної
структури беруть участь ферменти ДНК-глікозилази числом близько 20. Вони специфічно дізнаються про пошкодження, зумовлені дезамінуванням, алкілуванням та іншими структурними перетвореннями основ. Такі модифіковані підстави видаляються. Виникають ділянки, позбавлені підстав, які репаруються, як із втратою пуринів. Якщо відновлення нормальної структури не здійснюється, наприклад у разі дезамінування азотистих основ, відбувається заміна одних пар комплементарних основ іншими пара Ц-Г може замінюватися парою Т-А і т.п. (Див. Розд. 3.4.2.3).
Утворення в полінуклеотидних ланцюгах під дією УФ-променів тімінових димерів (Т-Т) вимагає участі ферментів, які впізнають не окремі змінені основи, а протяжні пошкодження структури ДНК. Репаративний процес у цьому випадку також пов'язаний з видаленням ділянки, що несе димер, та відновленням нормальної послідовності нуклеотидів шляхом синтезу на комплементарному ланцюзі ДНК.
У тому випадку, коли система ексцизійної репарації не виправляє зміни, що виникли в одному ланцюзі ДНК, під час реплікації відбувається фіксація
цієї зміни і воно стає надбанням обох ланцюгів ДНК. Це призводить до заміни однієї пари комплементарних нуклеотидів на іншу або до появи розривів (проломів) у знову синтезованому ланцюгу проти змінених ділянок. Відновлення нормальної структури ДНК може відбутися і після реплікації.
Постеплікативна репараціяздійснюється шляхом рекомбінації (обміну фрагментами) між двома новоствореними подвійними спіралями ДНК. Прикладом такої постреплікативної репарації може бути відновлення нормальної структури ДНК у разі тімінових димерів (Т-Т), коли де вони усуваються мимовільно під впливом видимого світла ( світлова репарація) або в ході дореплікативної ексцизійної репарації.
Ковалентні зв'язки, що виникають між залишками тиміну, що стоять поруч, роблять їх не здатними до зв'язування з комплементарними нуклеотидами. У результаті знову синтезованої ланцюга ДНК виникають розриви (проломи), відомі ферментами репарації. Відновлення цілісності нового полінуклеотидного ланцюга однієї з дочірніх ДНК здійснюється завдяки рекомбінації з відповідним їй нормальним материнським ланцюгом іншої дочірньої ДНК. Пробіл, що утворився в материнському ланцюгу, заповнюється потім шляхом синтезу на комплементарному їй полінуклеотидному ланцюгу (рис. 3.16). Проявом такої постреплікативної репарації, що здійснюється шляхом рекомбінації між ланцюгами двох дочірніх молекул ДНК, можна вважати обмін матеріалом між сестринськими хроматидами, що нерідко спостерігається (рис. 3.17).

Мал. 3.16. Схема постреплікативної репарації ДНК: I - виникнення тімінового димеру в одному з ланцюгів ДНК;
II - утворення «пролому» у знову синтезованому ланцюгу проти зміненої ділянки материнської молекули після реплікації (стрілкою показано наступне заповнення «пролому» ділянкою з відповідного ланцюга другої дочірньої молекули ДНК);
III - відновлення цілісності дочірнього ланцюга верхньої молекули за рахунок рекомбінації та в нижній молекулі за рахунок синтезу на комплементарному ланцюзі

Мал. 3.17. Міжхроматидні обміни (зазначені стрілками)
У ході дореплікативної та постреплікативної репарації відновлюється більшість пошкоджень структури ДНК. Однак, якщо в спадковому матеріалі клітини виникає занадто багато пошкоджень і частина з них не ліквідується, включається система індукованих ферментів репарації (SOS-система). Ці ферменти заповнюють проломи, відновлюючи цілісність полінуклеотидних ланцюгів, що синтезуються, без точного дотримання принципу комплементарності. Ось чому іноді самі процеси репарації можуть бути джерелом стійких змін у структурі ДНК (мутацій). Названа реакція відноситься до SOS-системі.
Якщо в клітині, незважаючи на репарацію, кількість пошкоджень структури ДНК залишається високою, в ній блокуються процеси реплікації ДНК. Така клітина не ділиться, а значить, не передає змін потомства, що виникли.
Зупинка клітинного циклу, що викликається пошкодженнями ДНК, у поєднанні з неможливістю молекулярної репарації зміненого спадкового матеріалу може за участю білка, синтез якого контролюється геном р53, призводити до активації процесу самоліквідації (апотпоз) дефектної клітини з метою усунення її з організму.
Таким чином, великий набір різних ферментів репарації здійснює безперервний огляд ДНК, видаляючи з неї пошкоджені ділянки і сприяючи підтримці стабільності спадкового матеріалу. Спільна дія ферментів реплікації (ДНК-полімераза і ендонуклеаза, що редагує) і ферментів репарації забезпечує досить низьку частоту помилок у молекулах ДНК, яка підтримується на рівні 1 · 10-9 пар змінених нуклеотидів на геном. При розмірі геному людини 3 · 109 нуклеотидних пар це означає поява близько 3 помилок на геном, що реплікується. Водночас, навіть цей рівень достатній для освіти за час існування життя на Землі значного генетичного розмаїття у вигляді генних мутацій.

3.4.2.3. Зміни нуклеотидних послідовностей ДНК. Генні мутації
Нескориговані зміни хімічної структуригенів, що відтворюються в послідовних циклах реплікації і проявляються у потомства у вигляді нових варіантів ознак, називають генними мутаціями.
Зміни структури ДНК, що утворює ген, можна поділити на три групи. Мутації першої групи полягають у заміні одних підстав іншими. Вони становлять близько 20% генних змін, що спонтанно виникають. Друга група мутацій обумовлена зсувом рамки зчитування, що відбувається за зміни кількості нуклеотидних пар у складі гена. Нарешті, третю групу являють собою мутації, пов'язані зі зміною порядку нуклеотидних послідовностей у межах гена (інверсії).
Мутації на кшталт заміни азотистих основ. Ці мутації відбуваються з низки конкретних причин. Однією з них може бути випадково або під впливом конкретних хімічних агентів зміна структури основи, вже включеної в спіраль ДНК. Якщо така змінена форма основи залишається не поміченою ферментами репарації, то при найближчому циклі реплікації вона може приєднувати інший нуклеотид. Прикладом може бути дезамінування цитозину, що перетворюється на урацил мимоволі або під впливом азотистої кислоти(Рис. 3.18). Урацил, що утворюється при цьому, не помічений ферментомДНК-глікозилаза,при реплікації з'єднується з аденіном, який згодом приєднує тимідиловий нуклеотид. В результаті параЦ-Г заміщається в ДНК пароюТ-А (рис. 3.19, І ). Дезамінування метильованого цитозину перетворює його на тімін (див. рис. 3.18). Тіміділовий нуклеотид, будучи природним компонентом ДНК, не виявляється ферментами репарації як зміна і при наступній реплікації приєднує аденіловий нуклеотид. В результаті замість париЦ-Г у молекулі ДНК також з'являється параТ-А (рис. 3.19, II).
Мал. 3.18. Спонтанне дезамінування цитозину
Іншою причиною заміни основ може бути помилкове включення до синтезованого ланцюга ДНК нуклеотиду, що несе хімічно змінену форму основи або його аналог. Якщо ця помилка залишається не поміченою ферментами реплікації та репарації, змінена основа входить у процес реплікації, що нерідко призводить до заміни однієї пари на іншу. Прикладом цього може бути приєднання в ході реплікації до аденіну материнського ланцюга нуклеотиду з 5-бромурацилом (5-БО), аналогічного тимідиловому нуклеотиду. При подальшій реплікації 5-БО охочіше приєднує не аденін, а гуанін. Гуанін у ході подальшого подвоєння утворює комплементарну пару з цитозином. В підсумку пара А-Тзамінюється у молекулі ДНК парою Г-Ц(Рис. 3.20).

Мал. 3. 19. Мутації за типом заміни основи (дезамінування азотистих основ у ланцюзі ДНК):
I - перетворення цитозину на урацил, заміна Ц-Г-пари на Т-А-пару;
II - перетворення метил-цитозину на тімін, заміна Ц-Г-пари на Т-А-пару
З наведених прикладів видно, що зміни структури молекули ДНК за типом заміни основ виникають або до, або в процесі реплікації спочатку одного полінуклеотидного ланцюга. Якщо такі зміни не виправляються під час репарації, то за наступної реплікації вони стають надбанням обох ланцюгів ДНК.
Мал. 3.20. Мутації за типом заміни основ
(Включення аналога азотистої основи при реплікації ДНК)
Наслідком заміни однієї пари комплементарних нуклеотидів на іншу є утворення нового триплету в нуклеотидній послідовності ДНК, що кодує послідовність амінокислот у пептидному ланцюзі. Це може й позначитися структурі пептиду у разі, якщо новий триплет буде «синонімом» колишнього, тобто. кодуватиме ту ж амінокислоту. Наприклад, амінокислота валін шифрується чотирма триплетами: ЦАА, ЦАГ, ЦАТ, ЦАЦ. Заміна третьої основи в будь-якому з цих триплетів не змінить його сенсу (виродженість генетичного коду).
У тому випадку, коли триплет, що знову виник, шифрує іншу амінокислоту, змінюються структура пептидного ланцюга і властивості відповідного білка. Залежно від характеру і місця заміни, що відбулася, специфічні властивості білка змінюються різною мірою. Відомі випадки, коли заміна лише однієї амінокислоти в пептиді істотно впливає на властивості білка, що проявляється у зміні складніших ознак. Прикладом може бути зміна властивостей гемоглобіну людини присерповидно-клітинної анемії (рис. 3.21). У такому гемоглобіні-(HbS) (на відміну від нормального НbА) - в р-глобінових ланцюгах у шостому положенні глутамінова кислота замінена валіном. Це є наслідком заміни однієї з основ у триплеті, що шифрує глутамінову кислоту (ЦТТ або ЦТЦ). Через війну з'являється триплет, шифруючий валін (ЦАТ чи ЦАЦ). В даному випадку заміна однієї амінокислоти в пептиді істотно змінює властивості глобіну, що входить до складу гемоглобіну (знижується його здатність зв'язуватися з 02), у людини розвиваються ознаки серповидноклітинної анемії.
У в деяких випадках заміна однієї підстави на іншу може призвести до появи одного знонсенс-триплетів (АТТ, АТЦ, АЦТ), що не шифрує жодної амінокислоти. Наслідком такої заміни буде переривання синтезу пептидного ланцюга. Підраховано, що заміни нуклеотидів в одному триплеті призводять у 25% випадків до утворення триплетів-синонімів; у 2-3-безглуздих триплетів, у 70-75%-до виникнення справжніх генних мутацій.
Таким чином, мутації за типом заміни основ можуть виникати як в результаті спонтанних змін структури основи в одному з ланцюгів вже існуючої подвійної спіралі ДНК, так і в ході реплікації у ланцюгу, що знову синтезується. У тому випадку, якщо ці зміни не виправляються в процесі репарації (або, навпаки, виникають у ході репарації), вони фіксуються в обох ланцюгах і відтворюватимуться далі в наступних циклах реплікації. Отже, важливим джерелом виникнення таких мутацій є порушення реплікації та репарації.
Мутації із зсувом рамки зчитування. Цей тип мутацій становить значну частку спонтанних мутацій. Вони відбуваються внаслідок випадання чи вставки в нуклеотидну послідовність ДНК однієї чи кількох пар комплементарних нуклеотидів. Більшість вивчених мутацій, що викликають
РЕПАРАЦІЯ ДНК
Системи репарації
2 Ексцизійна репарація. Приклади та види
3 Репарація помилок реплікації ДНК
4 Рекомбінантна (постреплікативна) репарація у бактерій
5 SOS-репарація
Системи репарації ДНК досить консервативні в еволюції від бактерій до людини та найбільш вивчені у Е. coli.
Відомі два типи репарації:пряма та ексцизійна
Пряма репарація
Пряма репарація - найпростіший шлях усунення пошкоджень у ДНК, у якому зазвичай задіяні специфічні ферменти, здатні швидко (зазвичай, одну стадію) усувати відповідне ушкодження, відновлюючи вихідну структуру нуклеотидів.
1. Так діє, наприклад,О6-метилгуанін-ДНК-метилтрансфераза
(фермент – самогубець), яка знімає мітильну групу з азотистої основи на один із власних залишків цистеїну
У Е. coli може за 1 хв синтезуватися до 100 молекул цього білка. Аналогічний за функціями білок вищих еукаріотів, очевидно, відіграє важливу роль у захисті від раку, що викликається внутрішніми та зовнішніми факторами алкілу.
ДНК-інсертаза
2. ДНК-інсертазами
фотоліаза
3. Тімінові димери "розшиваються" шляхом прямий репарацциза участюфотоліаз, що здійснюють відповідне фотохімічне перетворення. ДНК-фотоліази є групою ферментів, що активуються світлом, з довжиною хвилі 300 - 600 нм (видима область), для чого в їх структурі є особливий світлочутливий центр.
Вони широко поширені в природі та виявлені у бактерій, дріжджів, комах, рептилій, земноводних та людини. Ці ферменти потребують різноманітних кофакторів (FADH, тетрагідрофолієва кислота та ін.), що беруть участь у фотохімічній активації ферменту. Фотоліаза Е. coli є білок з молекулярною масою 35 кДа, міцно пов'язаний з олігорибонуклеотидом довжиною 10-15 нуклеотидівнеобхідним для активності ферменту.
Приклади прямої репарації
1. Метильована основа O 6- mG диметилюється ферментом метилтрансферазуО6-метилгуанін-ДНК-метилтрансфераза (фермент – самогубець), яка переносить мітильну групу на один із своїх залишків
цистеїну
2. АР-сайти можуть репаруватися шляхом прямої вставки пуринів за участю ферментів, які називаютьсяДНК-інсертазами(Від англ. insert-вставляти).
СХЕМА ПРИКЛАДУ ПРЯМОЇ РЕПАРАЦІЇ ПОШКОДЖЕНЬ У ДНК – метильована основа O6- mGдемітується ферментом метилтрансферазою, який переносить метильну групу на один зі своїх залишків амінокислоти цистеїну.
3. Фотоліаза приєднується до тімінового димеру і після опромінення цього комплексу видимим світлом (300-600 нм) димер розшивається.
СХЕМА ПРИКЛАДУ ПРЯМОЇ РЕПАРАЦІЇ ПОШКОДЖЕНЬ У ДНК – Фотоліаза
приєднується до тімінового димеру і після опромінення видимим спектром світла цей димер розшивається.

Ексцизійна репарація
(Від англ. excision - Вирізання).
ВИЗНАЧЕННЯ
Ексцизійна репарація включає видаленняпошкоджених азотистих основ з ДНК та подальше відновленнянормальної структури молекули.
МЕХАНІЗМ
В ексцизійній репарації зазвичай беруть участь кілька ферментів, а сам процес зачіпає
не тільки пошкоджений ,
але й сусідні з ним нуклеотиди .
УМОВИ
Для ексцизійної репарації необхідний другий (комплементарний) ланцюг ДНК. Загальна спрощена схема ексцизійної репарації представлена рис. 171.
ЕТАПИ
Першим етапом ексцизійної репарації є вирізування аномальних азотистих основ. Його каталізують групаДНК-N-Глікозилаз- ферменти, що розщеплюють глікозидний зв'язок між дезоксирибозою та азотистою основою.
ВАЖЛИВЕ ЗАУВАЖЕННЯ:
УлюдиниДНК-N-глікозилазимають високу субстратну специфічність: різні ферменти цього сімейства розпізнають і вирізають різні аномальні основи(8-оксогуанін, урацил, метилпурини та ін).
УбактерійДНК-N-глікозилазитакої субстратної специфічності не має
ЗАГАЛЬНІ ФЕРМЕНТИ ЕКСЦИЗІЙНОЇ РЕПАРАЦІЇ
| НАЗВА | ФУНКЦІЯ | МЕХАНІЗМ |
| ДНК-N-глікозилази | вирізання аномальних азотистих основ | розщеплює глікозидний зв'язок між дезоксирибозою та азотистою основою |
| АР-ендонуклеаза | створює умови для роботи наступного ферменту - екзонуклеази | розриває цукро-фосфатний кістяк молекули ДНК в АР-сайті |
| екзонуклеаза | вищеплює кілька нуклеотидів | послідовно відщеплює кілька нуклеотидів від пошкодженої ділянки одного ланцюга ДНК |
КОНКРЕТНІ НАСЛІДНІ КРОКИ ЦЬОГО МЕХАНІЗУ:
Внаслідок дії ДНК- N-Глікозилазутворюється АР-сайт, який атакується ферментом АР-ендонуклеазою. Вона розриває цукро-фосфатний кістяк молекули ДНК в АР-сайті і тим самим створює умови для роботи наступного ферменту. екзонуклеазияка послідовно відщеплює кілька нуклеотидів від пошкодженої ділянки одного ланцюга ДНК
У клітинах бактерій звільнене місце заповнюється відповідними нуклеотидами за участю ДНК-полімерази I, що орієнтується на другий (комплементарний) ланцюг ДНК.
Оскільки ДНК-полімераза I здатна подовжувати З"-кінець одного з ланцюгів у місці розриву в дволанцюгової ДНК і видаляти нуклеотиди з 5"-кінця того ж розриву,
тобто. здійснювати "нік-трансляцію" , цей фермент відіграє ключову роль у репарації ДНК. Остаточне зшивання репарованих ділянок здійснює ДНК-лігаза.
У клітинах еукаріотів (ссавців)
Ексцизійна репарація ДНК у клітинах ссавців супроводжується різким сплеском активності ще одного ферменту.полі АDР-рибозо-полімерази . При цьому відбувається ADP-рибозилювання білків хроматину(гістонів та негістонових білків), що веде до послаблення їх зв'язку з ДНК та відкриває доступ ферментам репарації.
Донором ADP-рибозиу цих реакціях виступаєNAD+, запаси якого сильно виснажуються при ексцизійній репарації ушкоджень, що викликаються рентгенівським опроміненням:

Негативно заряджені залишки ADP-рибозиіз внутрішнього складу молекули NAD+приєднуються через радикалглутаміновийкислоти або фосфосеринудо білків хроматину, що веде до нейтралізації позитивних зарядів цих білків та послаблення їхнього контакту з ДНК.
ЩО ПРЕДСТАВЛЯЄ СЕБЕ ГРУПА ФЕРМЕНТІВ
ДНК – глікозилази
розщеплює глікозидний зв'язок між дезоксирибозою та азотистою основою
що призводить до вирізування аномальних азотистих основ
ДНК – глікозилази , що беруть участь у усуненні окисних ушкоджень ДНК у клітинах прокаріот та еукаріот, Дуже різноманітні і відрізняються за субстратною специфічністю, просторовою структурою та способами взаємодії з ДНК.
До найбільш вивчених ДНК-глікозилазів відносяться:
ендонуклеаза III(EndoIII),
форм амідо піримідин-ДНК-глікозилазу (Fpg),
Mut Tі
Mut Yкишкової палички.
Ендонуклеаза IIIЕ. coli "дізнається" і специфічно вищеплює з ДНК окислені піримідинові основи.
Цей фермент являє собою мономерний глобулярний білок, що складається з 211 амінокислотнихзалишків (мол. маса 23,4 кДа). Ген, що кодує Endo III, секвенований, встановлена його нуклеотидна послідовність. Endo III є залізосерний білок [(4 Fe -4 S )2+-білок], що володіє елементом над вторинної структури типу "грецький ключ" (спіраль - шпилька - спіраль), службовцям для зв'язування з ДНК. Ферменти з аналогічною субстратною специфічністю та подібною амінокислотною послідовністю виділені також з клітин бика та людини.
Форм амідо піридин-ДНК-глікозилазу Е. coli "дізнається" і вищеплює з ДНК окислені гетероциклічні основи пуринового ряду .
СХЕМА ЕКЦИЗІЙНОЇ РЕПАРАЦІЇ СТАДІЯ 1
ДНКN

СХЕМА ЕКЦИЗІЙНОЇ РЕПАРАЦІЇ
1 ДНКNглікозидаза видаляє пошкоджену основу
АР ендонуклеазу вносить розрив у ДНК
2 Екзонуклеаза видаляє ряд нуклеотидів
3 ДНК полімераза заповнює звільнену ділянку комплементарними
мононуклеотидами
ДНК лігаза зшиває репарований ланцюг ДНК

Mut T- невеликий білок з молекулярною масою 15 кДа, що має нуклеозидтрифосфатазну активність, який переважно гідролізує dGTP до dGMP та пірофосфату.
Біологічна роль Mut T полягає у запобіганні освіті під час реплікації неканонічних парА:Gі А: 8-oxo-G.
Такі пари можуть з'являтися у разі, коли окислена форма
dGTP (8-oxo-dGTP)стає субстратомДНК-полімерази.
Mut Tгідролізує 8-oxo-dGTPу 10 разів швидше, ніж dGTP.
Це робить 8-oxo-dGTPнайкращим субстратомMutTта пояснює його функціональну роль.
Mut Yявляє собою специфічну аденін-ДНК-глікозилазу, що розщеплює N-глікозидний зв'язок між аденіном та дезоксирибозою. аденозину,утворює неканонічну пару з гуаніном.
Функціональна роль цього ферменту полягає у запобіганні мутації
T:A - G:A шляхом відщеплення непошкодженого залишку аденіназ пари основ A: 8-oxo-G.
Нуклеотидна ексцизійна репарація
(АТФ-залежний механізм видалення пошкоджень із ДНК)
Останнім часом в ексцизійній репарації особливу увагу приділяють АТР-залежний механізм видалення пошкоджень з ДНК. Цей вид ексцизійної репарації отримав назву нуклеотидна ексцизійна репарація (Nucleotide excision repair; NER).
Вона включає ДВА ЕТАПУ :
1. видалення з ДНКолігонуклеотидних фрагментів , що містять ушкодження, та
Ексінуклеаза
2. подальшу реконструкцію ланцюга ДНК за участю комплексу ферментів (нуклеаз, ДНК-полімерази, ДНК-лігази та ін).
Видалення фрагмента ДНК відбувається по обидва боки пошкодженогонуклеотиду. Довжина олігонуклеотидних фрагментів, що видаляються, відрізняється у прокаріотів і еукаріотів.
Видалення фрагмента ДНК у прокаріотів
Так, у Е. coli, Ст subtilus, Micrococcus luteus вирізається фрагмент довжиною 12-13 нуклеотидів,
Видалення фрагмента ДНК у еукаріотів
а у дріжджів, земноводних та людини - фрагмент, що складається з 24-32 нуклеотидів.
Ексінуклеаза- Фермент, що видаляє фрагменти ДНК
Вищеплення фрагмента ДНК здійснюється ферментомексінуклеазою(Excinuclease). У Е. coli цей фермент складається з 3 різних протомерів -
uvrA
uvr В
uvr З
кожен із яких виконує певну функцію під час ексцизійного вищеплення фрагмента ДНК. Назва цих білків дано за першими буквами слів"ultra violet repair".
Протомір uvr Амає АТРазну активність, зв'язується з ДНК у вигляді димера, здійснюючи
первинне розпізнавання пошкодження та
зв'язуванняuvr В
Протомір uvr Вмає:
1 . Латентною АТР-азною та латентною хеліказною активністю, необхідною для зміни конформацій та розплітання подвійної спіралі ДНК;
2. Ендонуклеазний активністю, розщеплюючи міжнуклеотидний (фосфодіефірний) зв'язок з бокуЗ"-кінцявищеплюваного фрагмента.
Протомір uvr Сдіє як ендонуклеаза, що вносить розрив у репарований ланцюг ДНК з5"-кінцяфрагмента, що вирізується.
Таким чином, протоміриuvr A, uvr, uvr Свзаємодіють із ДНК у певній послідовності, здійснюючи АТР-залежну реакціювищеплення олігонуклеотидного фрагмента з репарованого ланцюга ДНК.
Пролом, що утворився, в молекулі ДНК реставрується за участю ДНК-полімерази I і ДНК-лігази. Модель ексцизійної репарації за участю перерахованих вище ферментів представлена на рис. 172.
Ексцизійні репарації у людини
Ексцизійні репарації у людини також мають АТФ – залежний характер і включаютьтри основні етапи :
впізнавання пошкодження,
подвійне розрізання ланцюга ДНК,
відновний синтез та
лігування репарованого ланцюга.
Однак, в ексцизійній репарації ДНК людини беруть участь
25 різних поліпептидів ,
16 з яких беруть участь у вищепленні олігонуклеотидного фрагмента, будучи протомірамиексінуклеази,
а решта 9 здійснюють синтез ділянки молекули, що репарується.
У репараційній системі ДНК у людини дуже істотну рольвиконують білки транскрипції -
РНК-полімераза II і
TF ПН- один із шести основних факторів транскрипції еукаріот.
Слід зазначити, що ексцизійна репарація у прокаріотів, як і у еукаріотів, залежить від функціонального стану ДНК:
транскрибована ДНК репарується швидше,
чим транскрипційно неактивна.
Цей феномен пояснюється такими факторами:
структурою хроматину,
гомологією ланцюгів ділянок ДНК, що транскрибуються,
ефектом ушкодження ланцюгів та його впливом на РНК-полімеразу.
ВАЖЛИВЕ ЗАУВАЖЕННЯ:
КОДИРУЮЧИЙ ЛАНЦЮГ ДНК (ланцюг зберігання інформації)
МАТРИЧНИЙ ЛАНЦЮГ ДНК (з нього відбувається списування інформації)
Відомо, що такі великі ушкодження, як утворення тімінових димерів, блокують транскрипцію як у бактерій, так і у людини, якщо вони відбуваються на матричного ланцюгаДНК (пошкодження на кодуючоюланцюги не впливаютьна транскрипційний комплекс). РНК-полімераза зупиняється у місці пошкодження ДНК та блокує роботу транскрипційного комплексу.
Транскрипційно-репараційний фактор зчеплення (TRCF) .
У Е. coli посилення репарації при транскрипції опосередковується одним спеціальним білком.транскрипційно-репараційним фактором зчеплення (TRCF) .
Цей білок сприяє :
1. від'єднання РНК-полімерази від ДНК
2. одночасно стимулює освіту комплексу білків,
Здійснюють репарацію пошкодженої ділянки.
Після закінчення репарації РНК-полімераза встає місце і транскрипція триває (див. рис.).
Отже загальна схемаексцизійної репарації
1. ДНК-N -глікозилаза видаляє пошкоджену основу
2. АР-ендонуклеаза вносить розрив у ланцюг ДНК
3. Екзонуклеаза видаляє низку нуклеотидів
4. ДНК-полімераза заповнює звільнену ділянку
Комплементарними нуклеотидами
5. ДНК лігаза зшиває репарований ланцюг ДНК
Репарація помилок реплікації ДНК
шляхом метилювання
Помилки парування азотистих основ під час реплікації ДНК відбуваються досить часто (у бактерій один раз на 10 тис. нуклеотидів), внаслідок якиху дочірній ланцюг ДНК включаються некомплементарні нуклеотиди материнського ланцюга нуклеотиди -місметчі(англ. mismatch н е відповідати).
Незважаючи на те щоДНК-полімераза Iпрокаріот має здатність до самокорекції, її зусилля щодо усунення помилково приєднаних нуклеотидів. іноді виявляються недостатніі тоді в ДНК залишаються деякі неправильні (некомплементарні) пари.
У цьому випадку репарація відбувається з використанням певної системи, пов'язаної зметилюванням ДНК . Дія цієї системи репарації полягає в тому, що після реплікації через певний час (кілька хвилин) ДНК піддається метилюванню.
У Є. coli метилюєтьсяв основному аденін з освітою
N6-метил-аденіну (N6-mA).
До цього моменту знову синтезована(Дочірня)ланцюг залишається неметильованим.
Якщо в такому ланцюгу є неспарені нуклеотиди, то він піддається репарації: Таким чиномметилювання мітить ДНК та
включає систему виправлення помилок реплікації.
У цій системі репарації дізнаються особливі структури:
послідовністьG-N6-mA-T-Сі наступназа нею деформація
у подвійній спіралі у місці відсутності комплементарності (рис. нижче).
Усунення неспарених нуклеотидів у напівметильованоюмолекулі ДНК бере участь досить складний комплекс ферментів репарації, який сканує поверхню молекули ДНК,вирізає ділянку дочірнього ланцюга , що вдається до місметчам, а потім створює умови для забудовування
його необхідними (комплементарними) нуклеотидами.
Різні компоненти цього комплексу мають різні активностінуклеазний,
хеліказний,
Атразний,
необхідними для внесення розривів у ДНК і вищеплення нуклеотидів, розплітання подвійної спіралі ДНК та енергетичного забезпечення руху комплексу вздовж репарованої частини молекули.
Подібний за структурою та функціями комплекс ферментів репарації виявлено і в людини.
Рекомбінантна (постреплікативна) репарація
У тих випадках, коли з тих чи інших причин вищерозглянуті системи репарації виявляються порушеними, у ланцюгах ДНК можуть утворюватися проломи (недорепаровані ділянки), що мають іноді дуже суттєві розміри, Що загрожує порушенням системи реплікації і може призвести до загибелі клітин.
У цьому випадку клітина може використовувати для репарації однієї молекули ДНК іншу отриману після реплікації молекулу ДНК, тобто залучити для цієї мети механізмрекомбінації.
У бактерій
У бактерій у рекомбінантній репарації бере участьбілок Rec А. Він зв'язується з одноланцюжковою ділянкою ДНК і залучає його до рекомбінації згомологічними ділянками неушкоджених ланцюгів іншої молекули ДНК .
В результаті і розірвана (що містить проломи), і непошкоджена ланцюга молекули ДНК, що репарується, виявляються спаренимиз неушкодженими комплементарними ділянками ДНК, що відкриває можливість репарації шляхом охарактеризованих вище систем.
При цьому можуть відбуватися вирізанняпевного фрагмента та
заповненняз його допомогою проломи в дефектному ланцюзі.
Прогалини і розриви в ланцюгах ДНК, що виникають при цьому, заповнюються за участюДНК-полімерази I та ДНК-лігази .
SOS-репарація
Існування цієї системи вперше постулював М. Радман у 1974 р. Він же дав назву цьому механізму, включивши до нього міжнародний сигнал лиха "SOS" (врятуйте наші душі).
Ця система включається тоді, коли пошкоджень у ДНК стає настільки багато, що загрожує життю клітини. У цьому випадку відбувається індукція активності різноманітної групи генів, що задіяні в різних клітинних процесах, пов'язаних з репарацією ДНК.
Включення тих чи інших генів, що визначаються кількістю пошкоджень у ДНК, призводить до різних за значимістю клітинних відповідей (починаючи зі стандартної репарації пошкодженихнуклеотидів та закінчуючи придушеннямклітинного поділу).
Найбільш вивченаSOS-репараціяу Е. coli, головними учасниками якої є білки, що кодуються генами Rec AіLex А.
Перший з них є поліфункціональнимбілок Rec A, що бере участь
в рекомбінації ДНК, а також
в регуляції транскрипції генів фага лямбда, що вражає Е. coli,
а другий (Білок Lex А)є репресоромтранскрипції великої групи генів, призначених для репарації ДНКбактерій. При його інгібуванні чи дозволі репарація активується.
Зв'язування Rec А з Lex Анаводить до розщеплення останньогоі відповідно до активації генів репарації.
У свою чергу, індукція SOS-системи бактерії служить дляфага лямбда сигналом небезпекиі призводить до того, що профаг перемикається з пасивного на активний (літичний) шляхіснування, викликаючи цим загибель клітини-господаря.
SOS-система репарації виявлена не тільки у бактерій, а й у тварин та людини.
Гени, задіяні в SOS-репарації ушкоджень ДНК
| Гени | Наслідки активації гена |
| uvr А, В, С, D | Репарація пошкоджень вторинної структури ДНК |
| Rec А | Стреплікативна репарація, індукції SOS-системи |
| lex А | Вимкнення SOS-системи |
| rec N, ruv | Репарація двониткових розривів |
| Забезпечення рекомбінаційної репарації |
|
| umu С, D | Мутагенез, спричинений змінами властивостей ДНК-полімерази |
| sul А | Пригнічення клітинного поділу |
Висновок
Виправлення пошкоджень у ДНК тісно пов'язане з іншими фундаментальними молекулярно-генетичними процесами: реплікацією, транскрипцією та рекомбінацією.Всі ці процеси виявляються переплетенимив загальну системувзаємодій, що обслуговується великою кількістю різноманітних білків, багато з яких є поліфункціональними молекулами, задіяними в контролі реалізації генетичної інформації у клітинах про- та еукаріотів. Водночас очевидно, що природа "не скупиться"на елементах контролю, створюючи найскладніші системи корекції тих пошкоджень у ДНК, які несуть небезпекудля організму та особливо для його потомства. З іншого боку, у тих випадках, коли репараційних можливостей недостатньо для збереження генетичного статусу організму, настає необхідність програмованої клітинної смерті –апоптоз..
СХЕМА НУКЛЕОТИДНОЇ ЕКСЦИЗІЙНОЇ РЕПАРАЦІЇ У E. COLIЗ УЧАСТЬЮ ЕКСІНУКЛЕАЗИ
1. ТРАНСКРИПЦІЙНО НЕЗАЛЕЖНИЙ МЕХАНІЗМ

2. ТРАНСКРИПЦІЙНО ЗАЛЕЖНИЙ МЕХАНІЗМ

3. ЗАГАЛЬНИЙ ЕТАП РЕПАРАЦІЇ

УМОВНІ ПОЗНАЧЕННЯ
А – білокuvr А
Б - білокuvr У
С - білокuvr З
маленький чорний трикутник – знак вказує на місце пошкодження
СХЕМА РЕПАРАЦІЇ, ПОВ'ЯЗАНА З МЕТИЛЮВАННЯМ ДНК

Репарацією
називають здатність молекули ДНК "виправляти" які у її ланцюгах зміни. У відновленні вихідної структури беруть участь не менше 20 білків: які впізнають змінені ділянки ДНК і видаляють їх з ланцюга, що відновлюють правильну послідовність нуклеотидів і зшивають відновлений фрагмент з рештою молекул ДНК.
Перелічені особливості хімічної структури та властивостей ДНК зумовлюють виконувані їй функції. ДНК записує, зберігає, відтворює генетичну інформацію, бере участь у процесах її реалізації між новими поколіннями клітин та організмів.
Рибонуклеїнові кислоти - РНК -
представлені різноманітними за розмірами, структурою та виконуваними функціями молекулами. Усі молекули РНК є копіями певних ділянок молекули ДНК і, крім уже зазначених відмінностей, виявляються коротшими за неї і складаються з одного ланцюга. Між окремими комплементарними один одному ділянками одного ланцюга РНК можливе парування на підставі (А з У, Г з Ц) та утворення спіральних ділянок. В результаті молекули набувають специфічної конформації.
Матрична, або інформаційна, РНК (мРНК, іРНК) синтезуються в ядрі під контролем ферменту РНК-полімерази комплементарно-інформаційним послідовностям ДНК, переносить цю інформацію на рибосоми, де стає матрицею для синтезу білкової молекули. Залежно від обсягу копіюваної інформації молекула мРНК може мати різну довжину і становить близько 5% усієї клітинної РНК.
Рибособна РНК (рРНК) синтезується в основному в ядерці, в області генів рРНК і представлена різноманітними молекулярної масимолекулами, що входять до складу великої та малої субчастинок рибосом. Перед рРНК припадає 85% всієї РНК клітини.
Транспортна РНК (тРНК) становить близько 10% клітинної РНК. Існує понад 40 видів тРНК. При реалізації генетичної інформації кожна тРНК приєднує певну амінокислоту та транспортує її до місця збирання поліпентиду. У еукаріотів тРНК складаються з 70-90 нуклеотидів.
Будова клітини
Типи клітинної організації.
Серед усього різноманіття організмів, що нині існують на Землі, виділяють дві групи: віруси і фаги, які не мають клітинної будови; решта організмів представлені різноманітними клітинними формами життя. Розрізняють два типи клітинної організації: прокаріотичний
і еукаріотичний (
см рис.1 ).
Клітини прокаріотичного типу влаштовані порівняно легко. Вони немає морфологічно відокремленого ядра, єдина хромосома утворена кільцеподібної ДНК і у цитоплазмі; мембранні органели відсутні (їх функцію виконують різні вп'ячування плазматичної мембрани); у цитоплазмі є численні дрібні рибосоми; мікротрубочки відсутні, тому цитоплазма нерухома, а вії та джгутики мають особливу структуру. До прокаріотів відносять бактерії.
Більшість сучасних живих організмів відноситься до одного з трьох царств - рослин, грибів або тварин, що об'єднуються в надцарство еукаріотів.
Залежно від кількості, з яких складаються організми, останні ділять на одноклітинні та багатоклітинні. Одноклітинні організми складаються з однієї єдиної клітини, яка виконує всі функції. Багато хто з цих клітин влаштований набагато складніше, ніж клітині багатоклітинного організму. Одноклітинними є всі прокаріоти, а також найпростіші, деякі зелені водорості та гриби.
Зміст теми "Генетичні елементи бактерій. Мутації у бактерій. Трансдукція.":1. Мігруючі генетичні елементи бактерій. Транспозони. Бактеріофаги, як мігруючі генетичні елементи.
2. Мутація. Мутації бактерій. Мутаген. Спонтанні мутації. Зворотні мутації (реверсії).
3. Індуковані мутації бактерій. Хімічний мутагенез. Радіаційний мутагенез. Типи мутацій.
4. Репарація ДНК бактерій. Системи репарації ДНК. Компенсація функцій порушених у результаті мутацій. Інтрагенна супресія. Екстрагенна супресія.
5. Перенесення бактеріальної ДНК. Кон'югація бактерій. F-фактор бактерій.
6. Трансформація мікробів. Стадія трансформації бактерії. Картування хромосом бактерій.
7. Трансдукція. Неспецифічна трансдукція. Специфічна трансдукція. Абортивна трансдукція. Феномен лізогенії.
8. Властивості бактерій. Неуспадковані зміни властивостей бактерій. S – колонії. R – колонії. M – колонії. D – колонії бактерій.
Репарація бактерій ДНК. Системи репарації ДНК. Компенсація функцій порушених у результаті мутацій. Інтрагенна супресія. Екстрагенна супресія.
У клітині є механізми, здатні повністю або частково відновлювати вихідну структуру зміненої ДНК. Мутації, спричинені радіацією, хімічними речовинамита іншими факторами, теоретично могли б призвести до вимирання бактеріальної популяції, якби остання була позбавлена здатності до репарації ДНК. Сукупність ферментів, що каталізують корекцію ушкоджень ДНК, об'єднують у так звані системи репарації, які принципово різняться за біохімічними механізмами «залікування» ушкоджень. Відомо три основні напрями корекції дефектів ДНК.
1. Безпосередня реверсія від пошкодженої ДНКдо вихідної структури, коли зміни до ДНКвиправляються за допомогою єдиної ферментативної реакції. Наприклад, видалення неправильно приєднаної метильної групи при шостому атомі кисню гуаніну за допомогою метилтрансферази; або розщеплення, що виникло в результаті опромінення тімінового димера за допомогою фотоліази ( рекомбінаційна репарація).
2. «Вирізання» ушкодженьз подальшим відновленням вихідної структури (ексцизійна репарація).
3. Активація спеціальних механізмів, що забезпечують виживання при пошкодженнях ДНК(Відновлення вихідної структури ДНКвнаслідок рекомбінації; корекція помилкового спарювання основ; трансляційний синтез на пошкодженій матриці ДНК). Ці механізми не завжди призводять до повного відновлення вихідної структури ДНК.
Компенсація функцій, порушених внаслідок мутацій
Первинна мутаціяможе бути компенсована вторинною мутацією, яка сталася всередині гена, що мутував (інтрагенно) або в іншому гені (екстрагенно). Зміни, які усувають прояви мутації, не виправляючи при цьому первинного порушення ДНК, називають супресією.
Інтрагенна супресіявикликана вторинною мутацією, що коригує ефекти первинної мутації. Наприклад, точкова мутація, що призводить до синтезу дефектного білка з втраченою біологічною активністю, може бути виправлена, якщо вторинна мутація точна приведе до кодування амінокислоти, що зберігає конфігурацію і активність білка. Точне відновлення вихідної структури гена називають істинною зворотною мутацією ( справжньою реверсією). Якщо ефект першої мутації компенсовано мутацією в іншій частині гена, такі мутації називають вторинними реверсіями.
Екстрагенна супресія- Пригнічення прояву мутації, що сталася в одному гені, внаслідок мутації в другому гені.