Биологична химия Лелевич Владимир Валерианович
Глава 25
Метаболизъм на метионин
Метионинът е незаменима аминокиселина. Метиловата група на метионина е подвижен едновъглероден фрагмент, използван за синтеза на редица съединения. Прехвърлянето на метиловата група на метионина към подходящия акцептор се нарича трансметилиране, което е от голямо метаболитно значение. Метиловата група в молекулата на метионина е тясно свързана със серния атом, така че активната форма на аминокиселината служи като директен донор на едновъглеродния фрагмент.
Фигура 25.1. Обмен на метионин.
Реакция на активиране на метионин
Активната форма на метионин е S-аденозилметионин (SAM), получен в резултат на добавяне на метионин към молекула на аденозин. Аденозинът се образува от хидролизата на АТФ. Тази реакция се катализира от ензима метионин аденозин трансфераза, който присъства във всички типове клетки. Тя е уникална за биологични системи, тъй като това е единствената реакция, при която се освобождават и трите фосфатни остатъка на АТФ. Отцепването на метиловата група от SAM и нейното прехвърляне към акцепторното съединение се катализира от ензими на метилтрансферазата. SAM се превръща в S-аденозилхомоцистеин (SAH) по време на реакцията.
Реакциите на метилиране играят важна роля в организма и протичат много интензивно. Те се използват за синтезиране на:
1. фосфатидилхолин от фосфатидилетаноламин;
2. карнитин;
3. креатин;
4. епинефрин от норепинефрин;
5. метилиране на азотни бази в нуклеотиди;
6. инактивиране на метаболити (хормони, медиатори) и неутрализиране на чужди съединения.
Всички тези реакции причиняват голяма консумация на метионин, тъй като той е незаменима аминокиселина. Поради тази причина играе голямо значениевъзможност за регенериране на метионин. В резултат на разцепването на метиловата група SAM се превръща в SAG, който под действието на хидролаза се разцепва на аденозин и хомоцистеин. Хомоцистеинът може да бъде превърнат обратно в метионин чрез хомоцистеин метилтрансфераза. Донорът на метиловата група в този случай е 5-метилтетрахидрофолиева киселина (5-метил-THFA), която се превръща в THFA. Междинният носител на метиловата група в тази реакция е производно на витамин В 12 - метилкобаламин, който действа като коензим. Доставчикът на едновъглеродни фрагменти за регенериране на 5-метил-THPA е серинът, който се превръща в глицин.
Синтез на креатин
Креатинът е необходим за образуването на макроергичното съединение креатин фосфат в мускулите. Синтезът на креатин се извършва на 2 етапа с помощта на 3 аминокиселини: аргинин, глицин и метионин. В бъбреците гуанидиноацетатът се образува под действието на глицин амидинотрансфераза. След това гуанидиноацетатът се транспортира до черния дроб, където протича реакцията му на метилиране с образуването на креатин. Креатинът се транспортира с кръвен поток до мускулите и мозъчните клетки, където се образува от него под действието на креатин киназата (реакцията е лесно обратима), за да се образува креатин фосфат – своеобразно енергийно депо.
Метаболизъм на фенилаланин и тирозин
Фенилаланинът е незаменима аминокиселина, тъй като неговият бензенов пръстен не се синтезира в животински клетки. Метаболизмът на метионина се осъществява по 2 начина: той се включва в протеините или се превръща в тирозин под действието на специфична монооксигеназа - фенилаланин хидроксилаза. Тази реакция е необратима и играе важна роля за отстраняването на излишния фенилаланин, тъй като високите концентрации от него са токсични за клетките.
Обменът на тирозин е много по-сложен. Освен че се използва в протеиновия синтез, тирозинът в различни тъкани действа като предшественик на такива съединения като катехоламини, тироксин, меланин и др.
В черния дроб тирозинът се катаболизира до крайните продукти фумарат и ацетоацетат. Фумаратът може да се окисли до CO 2 и H 2 O или да се използва за глюконеогенеза.
Преобразуване на тирозин в меланоцити. Той е предшественик на меланините. Синтезът на меланини е сложен многоетапен процес, като първата реакция - превръщането на тирозин в DOPA - се катализира от тирозиназа, която използва медни йони като кофактор.
Щитовидната жлеза синтезира хормоните тироксин и трийодтиронин от тирозин.
В надбъбречната медула и нервната тъкан тирозинът е предшественик на катехоламините. Междинният продукт от техния синтез е DOPA. Въпреки това, за разлика от меланоцитите, хидроксилирането на тирозин се осъществява под действието на тирозин хидроксилаза, която е Fe 2+-зависим ензим и неговата активност регулира скоростта на синтеза на катехоламини.
От книга Разказбиология [От алхимията до генетиката] автор Азимов ИсакГлава 12 Метаболизъм Химиотерапия Борбата с бактериални заболявания е много по-лесна от борбата с вирусни заболявания. Както вече беше показано, бактериите се размножават по-лесно в културата. Бактериите са по-уязвими. Живеейки извън клетката, те причиняват увреждане на тялото, отнемайки храната от него или
От книгата Фармацевтична и хранителна мафия от Брауър ЛуисКонцентрацията на лекарите в отделни региони Ако анализираме картата, характеризираща гъстотата на лекарите в частна практика, можем да получим следната представа за 👉🏻♀️ в Южна Франция има 360 жители на лекар; на север
От книгата ПЕЧАТ НА СЪЗДАТЕЛЯ. Хипотеза за произхода на живота на Земята. автор Филатов Феликс ПетровичИндивидуални лабораторни практики Пример са търговските методи, използвани от Siba Geigy Laboratories, които произвеждат не само лекарства, но и продукти, предназначени за селското стопанство, по-специално пестициди. Тези лаборатории
От книгата на фенетиката [Еволюция, население, знак] автор Яблоков Алексей ВладимировичМафиотски практики на отделни лаборатории Според разследване на Мишел Редиа, мултинационалното общество Merek, със седалище в щата Ню Джърси в Съединените щати, започва да се занимава с измамни дейности. Измама
От книгата Гени и развитие на тялото автор Нейфах Александър АлександровичГлава 496 (XII) На неопитния читател може да изглежда, че елементите на генетичната кодираща машина са описани в предишната глава толкова подробно, че до края на четенето той дори започва да се уморява някак си, чувствайки, че няколко
От книгата Марката на създателя автор Филатов Феликс ПетровичПринципът на индивидуалните черти Г. Мендел имаше един невероятен предшественик, който почти разкри основните закони за унаследяване на черти 35 години преди работата на Мендел. Това е О. Сажре - френски селекционер (1763-1851), автор на забележителни трудове за хибридизацията на тиквата,
От книгата Биологична химия автор Лелевич Владимир ВалериановичОт индивидуални знаци до сешоар Английският биолог У. Батсън беше един от тези, които формулираха цели научни направления, прозорливо разграничаващи „съществено от несъществено“. Той предлага термина "генетика" през 1906 г. и е организатор на първите конференции по хибридизация,
От книгата на автора3. Движение на отделни клетки Много процеси на морфогенеза протичат не чрез движение на клетъчните слоеве, а чрез движение на отделни клетки. Понякога движението на клетъчния слой се случва с частична загуба на епителната му структура. Например,
От книгата на автораГлава 496 (XII) На неопитния читател може да изглежда, че елементите на генетичната кодираща машина са описани в предишната глава толкова подробно, че до края на четенето той дори започва да се уморява някак си, чувствайки, че няколко
От книгата на автораГлава 8 Въведение в метаболизма химична реакцияв тялото, които му осигуряват необходимите за живота вещества и енергия. Метаболитният процес, който е придружен от образуването на по-прости
От книгата на автораМетаболизъм на фруктоза Значително количество фруктоза, което се образува по време на разграждането на захарозата, се превръща в глюкоза още в чревните клетки, преди да влезе в системата на порталната вена. Друга част от фруктозата се усвоява с помощта на протеин-носител, т.е. през
От книгата на автораМетаболизъм на галактозата Галактозата се образува в червата в резултат на хидролизата на лактозата.Нарушението на метаболизма на галактозата се проявява при наследствено заболяване - галактоземия. Това е резултат от вроден дефект на ензима
От книгата на автораМетаболизъм на лактозата Лактозата, дизахарид, открит само в млякото, се състои от галактоза и глюкоза. Лактозата се синтезира само от секреторните клетки на жлезите на бозайниците по време на кърмене. Присъства в млякото в количество от 2% до 6% в зависимост от вида.
От книгата на автораГлава 22 Биохимия на атеросклерозата Холестеролът е стероид, който се среща само в животински организми. Основното място на образуването му в човешкото тяло е черният дроб, където се синтезира 50% от холестерола, 15-20% се образува в тънките черва, останалата част
От книгата на автораГлава 23 Динамичното състояние на телесните протеини Значението на аминокиселините за организма се крие преди всичко във факта, че те се използват за синтеза на протеини, чийто метаболизъм заема специално място в процесите на метаболизъм между организма и
От книгата на автораМетаболизъм на метионин Метионинът е незаменима аминокиселина. Метиловата група на метионина е подвижен едновъглероден фрагмент, използван за синтеза на редица съединения. Прехвърлянето на метиловата група на метионина към подходящия акцептор се нарича трансметилиране,
Тема 1. РОЛЯТА НА ПРОТЕИНИТЕ В ХРАНЕНИЕТО. СМИРАНЕ НА ПРОТЕИН
Практическата значимост на темата.Хранителните протеини са основният и основен източник на аминокиселини за хората. За задоволяване на необходимите нужди на организма от аминокиселини са важни не само състоянието на стомашно-чревния тракт и количеството протеини в храната, но и качествените характеристики на хранителните протеини. Колкото по-близък е аминокиселинният състав на хранителния протеин до аминокиселинния състав на телесните протеини, толкова по-висока е степента на неговата абсорбция в храносмилателния тракт. Освен това тъканните протеини на тялото, които постоянно се метаболизират с освобождаването на съставните им мономери, могат да служат като източници на аминокиселини.
Целта на урока.След изучаване на тази тема студентът трябва да познава условията, необходими за ефективното усвояване на протеините и усвояването на аминокиселините в стомашно-чревния тракт, факторите, които допринасят за гниенето на аминокиселините в червата и механизмите за неутрализиране на продуктите на гниене и тяхната биологична роля, да могат да прилагат получените знания за решаване на теоретични и практически проблеми.
Първоначално ниво на знания
- Структурата на аминокиселините (аланин, аспартат, глутамат, хистидин, тирозин, триптофан, цистеин).
- Коензимни функции на витамини (пиридоксин, никотинамид).
- Хидрофилни и хидрофобни свойства на органичните съединения.
- Свойства на ензимите поради тяхната химическа природа.
- Механизми за регулиране на активността на каталитичните протеини.
- Редокс реакции, реакции на трансфер и елиминиране на функционални групи в биологичните системи.
Аминокиселинен фонд на тялото.
1.1.1. Човешкото тяло съдържа около 100 g свободни аминокиселини, които образуват неговия аминокиселинен фонд. Този фонд непрекъснато се попълва поради получаването на нови молекули на аминокиселини, които да заменят тези, които са били използвани в метаболитните процеси. Източниците и начините за използване на свободните аминокиселини в организма са показани на Фигура 1.1.
Фигура 1.1.Образуване и използване на свободни аминокиселини в организма.
1.1.2. Изследвания с използване на радиоактивни етикети показват, че при здрав възрастен, общата скорост на протеинов синтез в тялото е около 400-500 g на ден, като 3/4 от този синтез се осигурява от ендогенни ресурси. Това обяснява факта, че дори по време на гладуване, синтезът на определени протеини се извършва с доста висока скорост.
азотен баланс.
1.2.1. За правилна оценка на съотношението на процесите на биосинтеза и разграждането на протеините в организма, доста точен параметър е азотният баланс. Азотният баланс е разликата между количеството азот, погълнат с храната, и количеството азот, отделен от тялото с урина, изпражнения, слюнка и пот.
1.2.2. Ако количеството поет азот надвишава количеството освободен азот, тогава положителен азотен баланс . Характерно е за всички състояния, при които скоростта на синтеза на протеин в тялото е по-висока от скоростта на неговото разпадане, например:
- при жени по време на бременност;
- в детска възраст с добро хранене;
- при пациенти по време на възстановителния период;
- при спортисти по време на тренировка;
- с въвеждането на анаболни хормони.
1.2.3. Ако количеството азот, отделен от тялото, надвишава количеството азот, доставен с храна, тогава отрицателен азотен баланс . Появява се във всички случаи, когато разграждането на протеините в организма надделя над синтеза им, например.
Предговор
Протеините са в основата на жизнената дейност на всички организми, познати на нашата планета. Това са сложни органични молекули, които имат голямо молекулно тегло и са биополимери, състоящи се от аминокиселини. Клетъчните биополимери включват и нуклеинови киселини – ДНК и РНК, които са резултат от полимеризацията на нуклеотидите.
протеинов метаболизъм и нуклеинова киселинавключва синтеза им от структурните компоненти на аминокиселините и съответно нуклеотидите и разпадането им до тези мономери, последвано от разграждането им до крайните продукти на катаболизма – CO 2 , H 2 O, NH 3 , пикочна киселина и др.
Тези процеси са химически сложни и практически няма алтернативни байпаси, които да функционират нормално в случай на метаболитни нарушения. Известни наследствени и придобити заболявания, чиято молекулярна основа са промените в метаболизма на аминокиселините и нуклеотидите. Някои от тях имат тежки клинични прояви, но, за съжаление, в момента не съществуват. ефективни методитяхното лечение. Говорим за заболявания като подагра, синдром на Lesch-Nyhan, ензимопатии на метаболизма на аминокиселините. В тази връзка подробното изследване на обмена на аминокиселини и нуклеотиди в нормата и техните възможни нарушения е от голямо значение за формирането на арсенал от теоретични знания, необходими в практиката на лекаря.
При написването на конспекта от лекцията „Метаболизъм на аминокиселини и нуклеотиди“ авторите не си поставят задачата да опишат подробно всички химични процеси и трансформации на аминокиселини и нуклеотиди, които любознателният студент може да намери във всеки учебник по биохимия. Основната задача беше да се представи материалът по такъв начин, че сложните биохимични реакции да се възприемат лесно, достъпно, разбираемо, с подчертаване на основното. За „силните“ студенти лекционните материали могат да станат отправна точка в последващо, по-задълбочено изследване на биохимичните трансформации. За тези, които не са се превърнали в любим предмет на биохимията, лекциите ще помогнат за формиране на основата на биохимичните знания, необходими при изучаването на клинични дисциплини. Авторите изразяват надежда, че предложените конспекти от лекции ще се превърнат в добър помощник на студентите по пътя към тяхното бъдеща професия.
Тема. Метаболизъм на аминокиселини: общи пътища на метаболизма. Синтез на урея
Планирайте
1 Начини за трансформация на аминокиселините в тъканите.
2 Трансаминиране на аминокиселини.
3 Дезаминиране на аминокиселини. Непряко дезаминиране.
5 Обмен на амоняк. биосинтеза на урея. Някои клинични аспекти.
1 Пътища за трансформация на аминокиселините в тъканите
Аминокиселините са основният източник на азот за бозайниците. Те са връзка между процесите на синтез и разлагане на азотсъдържащи вещества, предимно протеини. На ден в човешкото тяло се обновяват до 400 g протеин. Като цяло периодът на разпадане на всички протеини на човешкото тяло е 80 дни. Четвъртата част от протеиновите аминокиселини (около 100 g) се разпада необратимо. Тази част се обновява благодарение на хранителните аминокиселини и ендогенния синтез – синтеза на неесенциални аминокиселини.
В клетките постоянно се поддържа определено стационарно ниво на аминокиселини – фонд (пул) от свободни аминокиселини. Този фонд се обновява поради приема на аминокиселини и се използва за синтеза на биологично важни химични компоненти на клетката, т.е. могат да бъдат идентифицирани пътища за влизане и използване клетъчен пул от аминокиселини.
Входни маршрутисвободни аминокиселини, които образуват групата от аминокиселини в клетката:
1 Транспорт на аминокиселини от извънклетъчната течност- транспортират се аминокиселини, които се абсорбират в червата след хидролизата на хранителните протеини.
2 Синтез на неесенциални аминокиселини- аминокиселините могат да се синтезират в клетката от междинните продукти на глюкозното окисление и цикъла на лимонената киселина. Неесенциалните аминокиселини включват: аланин, аспарагинова киселина, аспарагин, глутаминова киселина, глутамин, пролин, глицин, серин.
Вътреклетъчна хидролиза на протеиние основният път за прием на аминокиселини. Хидролитичното разцепване на тъканните протеини се катализира от лизозомни протеази. При гладуване, онкологични и инфекциозни заболявания този процес се засилва.
Начини за използванефонд от аминокиселини:
1) Синтез на протеини и пептиди- това е основният начин на консумация на аминокиселини - 75-80% от аминокиселините в клетката отиват за техния синтез.
2) Синтез на непротеинови азотсъдържащи съединения:
Пуринови и пиримидинови нуклеотиди;
порфирини;
креатин;
меланин;
Някои витамини и коензими (NAD, CoA, фолиева киселина);
Биогенни амини (хистамин, серотонин);
Хормони (адреналин, тироксин, трийодтиронин);
Медиатори (норепинефрин, ацетилхолин, GABA).
3) Синтез на глюкозаизползване на въглеродни скелети от гликогенни аминокиселини (глюконеогенеза).
4) В липиден синтезизползвайки ацетилови остатъци от въглеродни скелети на кетогенни аминокиселини.
5) Окисление до крайни продукти на метаболизма (CO 2 , H 2 O, NH 3) е един от начините за осигуряване на клетката с енергия – до 10% от общите енергийни нужди. Всички аминокиселини, които не се използват в синтеза на протеини и други физиологично важни съединения, се разцепват.
Има общи и специфични пътища на метаболизма на аминокиселините. Общите пътища на катаболизъм за аминокиселини включват:
1) трансаминиране;
2) дезаминиране;
декарбоксилиране.
2 Трансаминиране на аминокиселини
трансаминиранеаминокиселини - основният начин за дезаминиране на аминокиселини, което протича без образуването на свободен NH3. Това е обратим процес на прехвърляне на NH2 група от аминокиселина към α-кето киселина. Процесът е открит от A.E. Браунщайн и М.Б. Крицман (1937).
Всички аминокиселини могат да участват в трансаминирането, с изключение на треонин, лизин, пролин и хидроксипролин.
Реакцията на трансаминиране в общ изгледкакто следва:
UNSD UNSD UNSD UNSD
HC - NH 2 + C \u003d O C \u003d O + HC - NH 2
R1 Р 2 R1 Р 2
аминокиселина
-кетокиселина
Ензимите, които катализират този тип реакции, се наричат аминотрансферази (трансаминаза-mi). Аминотрансферазите на L-аминокиселините функционират в човешкото тяло. Акцептор на аминогрупата в реакцията са -кето киселини – пируват, оксалоацетат, -кето-глутарат. Най-често срещаните аминотрансферази са ALT (аланин аминотрансфераза), AST (аспартат аминотрансфераза), тирозин аминотрансфераза.
Реакцията, катализирана от ензима ALT, е показана по-долу:
UNSD UNSD UNSD UNSD
│ │ AlAT│ │
HCNH 2 + C \u003d O C \u003d O + HCNH 2
│ │ │ │
CH 3 CH 2 CH 3 CH 2
Ала │ PVC │
- кетоглутарат Дълбок
Реакцията, катализирана от ензима AST, може да бъде схематично изобразена, както следва:
Asp + -кетоглутарат Оксалоацетат + Glu.
Коензимни трансаминази- пиридоксал фосфат (В 6) - е част от активния център на ензима. В процеса на трансаминиране коензимът действа като носител на аминогрупа и двете коензимни форми на PALF (пиридоксал-5-f) и PAMF (пиридоксамин-5-f) се преобразуват взаимно:
NH2 група
PALF PAMF.
NH2 група
Трансаминирането протича активно в черния дроб. Това ви позволява да регулирате концентрацията на всякакви аминокиселини в кръвта, включително тези, получени с храна (с изключение на tre, lis, pro). Благодарение на това оптималната смес от аминокиселини се пренася с кръвта до всички органи.
В някои случаи може да възникне нарушение на трансаминирането на аминокиселини:
1) с хиповитаминоза B 6;
2) при лечение на туберкулоза с антагонисти на трансами-аз - фтивазид и неговите аналози;
3) при гладуване, цироза и стеатоза на черния дроб има липса на синтез на протеиновата част на трансаминазите.
За диагнозата е важно определянето на активността на аминотрансферазите в кръвната плазма. При патологични състояния се наблюдава увеличаване на цитолизата в определен орган, което е придружено от повишаване на активността на тези ензими в кръвта.
Отделни трансаминази се намират в различни тъкани в различни количества. AsAT е повече в кардиомиоцитите, черния дроб, скелетните мускули, бъбреците, панкреаса. AlAT - в рекордно количество в черния дроб, в по-малка степен - в панкреаса, миокарда, скелетните мускули. Следователно повишаването на активността на AST в кръвта е по-характерно за инфаркт на миокарда (MI), а повишаването на активността на ALT може да показва цитолиза в хепатоцитите. Така при остър инфекциозен хепатит в кръвта активността на AlAT > AsAT; но с цироза на черния дроб - AsAT > AlAT. Леко повишаване на активността на ALT се наблюдава и при МИ. Следователно, определянето на активността на две трансаминази наведнъж е важен диагностичен тест. Обикновено съотношението на активността на AST/ALAT (коефициент на де Ритис) е 1,330,42. При МИ стойността на този коефициент се увеличава рязко, при пациенти с инфекциозен хепатит, напротив, този показател намалява.
3 Дезаминиране на аминокиселини. Непряко разминиране
Процесът е тясно свързан с трансаминирането окислително дезаминиране, в резултат на което NH 2 групата се отцепва с образуването на NH 3, H 2 O и -кето киселини. Деаминирането на аминокиселините се извършва най-активно в черния дроб и бъбреците.
Процесът се катализира от ензими оксидаза,които са флавопротеини. Има оксидази на L- и D-аминокиселини. Оксидази на L-аминокиселини FMN-зависими, D-аминокиселини FAD-зависими.
Реакцията на окислително деаминиране на L-аминокиселини може да бъде схематично представена, както следва:
FMD FMN N + H 2 O NH 3
Л– АКЛ– иминокиселина -кетокиселини.
В човешкото тяло активността на аминокиселинните оксидази е изключително ниска.
Окислителното дезаминиране на L-глутаминова киселина протича най-активно в клетките:
НАД НАДН H + H 2 O
L-глутамат L-иминоглутарат -KG + NH 3 .
1
2
1
- Глутамат дехидрогеназа(може да използва както NAD +, така и NADP +);
2 - Този етап е неензимно.
Схематично общо уравнениереакции (тази реакция е обратима):
L-Glu + NAD + H 2 O -KG + NADH H + + NH 3
Л– глутамат дехидрогеназа- ензим, който катализира тази реакция, който има висока активност и е широко разпространен в тъканите на бозайниците.
Чернодробната глутамат дехидрогеназа е регулаторен ензим, локализиран в митохондриите. Активността на този ензим зависи от енергийното състояние на клетката. При енергиен дефицит реакцията протича в посока на образуване на -кетоглутарат и NADH. H + , които се изпращат съответно към CLA и окислително фосфорилиране. В резултат на това се наблюдава увеличаване на синтеза на АТФ в клетката. Следователно за глутамат дехидрогеназата инхибиторите са ATP, GTP, NADH, а активаторът е ADP.
Повечето аминокиселини се деаминират от непряко дезаминиранее процесът на конюгиране на 2 реакции:
1 ) трансаминиране с образуване на глутамат;
2
)
глутамат дехидрогеназна реакция.
аминокиселина -KG NADH H +
NH3 1
2
NH3
-кето киселина глутамат NAD
В този случай биологичното значение на трансаминирането ( 1
)
се състои в събиране на аминогрупите на всички разлагащи се аминокиселини под формата на аминокиселина от един вид - глутамат. Освен това глутаминовата киселина се транспортира до митохондриите, където се подлага на окислително деаминиране под действието на глутамат дехидрогеназа ( 2
).
Най-активното непряко дезаминиране се извършва в черния дроб. Тук полученият NH 3 влиза в цикъла на урея за неутрализиране.
Посоката на равновесните процеси на трансаминиране, индиректно дезаминиране до голяма степен зависи от наличието и концентрацията на аминокиселини и -кето киселини. При излишък от аминоазот се засилва превръщането на аминокиселините в съответните кетокиселини, последвано от тяхното енергийно и пластично използване.
4 Декарбоксилиране на аминокиселини
Това е процесът на разцепване на карбоксилната група, която се намира в -позиция на аминокиселината, с образуването на амини и CO 2. В резултат на декарбоксилирането на аминокиселините се образуват:
биогенни амини (хистамин, допамин, тирамин, -аминомаслена киселина - GABA и др.).
UNSD CH2NH2
CHNH 2 ТАКА 2 CH 2
CH2COOH
Glu GABA
Декарбоксилирането на аминокиселини с образуването на биогенни амини протича най-активно в черния дроб, мозъка и хромафинната тъкан.
2) продукти на "гниещи протеини в червата", които са резултат от декарбоксилиране на аминокиселини под въздействието на чревната микрофлора. Аминокиселините образуват токсични продукти, например:
-CO 2
лизин кадаверин
-CO 2
орнитин путресцин
Общо в човешкото тяло се образуват повече от 40 различни амини. По време на хипоксия и гладуване се наблюдава увеличаване на синтеза на амини. Локално повишаване на синтеза, освобождаването и инактивирането на катехоламини, хистамин и серотонин е характерно за огнища на възпаление.
Злокачествените тумори с апудоцитен произход, разположени в червата, бронхите, панкреаса, могат да синтезират голямо количество серотонин (използвайки за тази цел до 60% от дневната нужда от триптофан).
Биогенните амини са инактивиранипод действието на окислителни FAD-зависими ензими - моноаминооксидази (МАО). Настъпва окислително дезаминиране на амини до алдехиди.
R–CH 2 –NH 2 + FAD + H 2 O R–CH + NH 3 + FADH 2
Продукти на деаминиране на биогенни амини - алдехиди– окислено към органични киселиникато се използва алдехид дехидрогенази.Тези киселини се екскретират с урината или се подлагат на по-нататъшно окислително разграждане. В допълнение, катехол-О-метилтрансфераза участва в разграждането на катехоламините.
Някои клинични аспекти
При условия на МАО блокада (по време на терапия с антидепресанти) способността за унищожаване на амините намалява. В този случай тялото може да стане чувствително към действието на амините. Например, ядене на сирене и пиене на определени сортове червено вино, които са богати на тирамин, на фона на терапия с МАО инхибитори води до хипертония.
Намаляване на активността на МАО се наблюдава при излишък от хормони на щитовидната жлеза.
Повишаване на активността на МАО може да възникне при авитаминоза B 1, tk. един от метаболитните продукти В 1 е МАО инхибитор.
5 Обмен на амоняк. биосинтеза на урея. Някои клинични аспекти
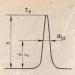
Амонякът е един от крайните продукти на метаболизма на азотсъдържащите вещества. Това е компонент от фракцията на остатъчния азот в кръвния серум (заедно с урея, пикочна киселина, креатинин, индикан). В кръвта концентрацията на амоняк е ниска - 25-40 µmol / l. При по-високи концентрации има токсичен ефект върху организма.
Амонякът е токсичен, главно за централната нервна система. Токсичността на амоняка е свързана с неговата способност да нарушава функционирането на CLC, tk. NH 3 премахва -кетоглутарата от CLA:
-KG + NH 3 + NADH. H + Glu + НАД + + H 2 O.
В крайна сметка редукционно аминиране-кето-глутарат има намаляване на активността на CLA в клетките на централната нервна система, което от своя страна инхибира активността на аеробното окисление на глюкоза. В резултат на това се нарушава производството на енергия и се развива хипоенергийно състояние. Глюкозата е основният източник на енергия за мозъка.
NH
3
образувани по време на следните процеси
:
1) окислително дезаминиране на аминокиселини – това е основният път за производството на NH 3;
дезаминиране на биогенни амини;
дезаминиране на пуринови основи (аденин, гуанин);
катаболизъм на пиримидинови нуклеотиди.
AMP + H 2 O IMP + NH 3.
Ензимът, който катализира тази реакция е аденозин дезаминаза.
Амонякът се транспортира чрез кръвта до черния дроб и бъбреците за неутрализиране в състава на аминокиселини, сред които основните са глутамин, аспарагин, аланин.
Неутрализацията на NH 3 настъпва почти веднага след образуването му, т.к. в тъканите веднага се включва в състава на аминокиселините, главно глутамин. Въпреки това, за по-нататъшна детоксикация и отделяне на амоняка, има биохимични процеси в черния дроб и бъбреците, които са основните начини за неутрализиране на NH 3.
Има следните механизми за неутрализиране NH 3 :
1 ) редуктивно аминиране на -кетоглутарат;
2 ) образуване на аминокиселинни амиди - аспарагин и глутамин;
3 ) образуване на амониеви соли в бъбреците;
4 ) синтез на урея.
В тъканите амонякът подлежи на незабавна неутрализация. Това се постига чрез комбинация от процеси ( 1 ) и ( 2 ).
Редукционно аминиране -кетоглутарат:
NH 3 + –KG + NADH . Х + Glu + NAD + H 2 О
Ензим - глутамат дехидрогеназа
Този процес изисква значителни концентрации на -KG. За да се избегне преразход на -KG и работата на CLC да не се нарушава, -KG се попълва поради трансформацията на PVC OA -KG.
2 ) Образуване на амиде важен спомагателен механизъм за детоксикация на NH 3 в тъканите чрез свързването му с Glu или Asp.
Asp + ATP +NH 3 Asn + AMF + FF nn
Ензим - аспарагин синтаза
Glu + ATP +NH 3 Gln + AMF + FF nn
ензим - глутамин синтаза
Този процес е най-активен в централната нервна система, мускулите, бъбреците, черния дроб (за поддържане на вътрешната концентрация на NH 3). Основно glnе транспортна форма на нетоксичен NH 3 от мозъка, мускулите и други тъкани. Глутаминът лесно прониква през мембраната, т.к. при физиологични стойности на pH няма заряд. По време на тренировка аланинът активно транспортира NH3 от мускулите към черния дроб. Освен това голямо количество аланин съдържа кръв, изтичаща от червата. Този аланин също се изпраща в черния дроб за глюконеогенеза.
3 ) Gln и asn с кръвния поток влизат в бъбреците, където се подлагат на хидролиза с помощта на специални ензими - глутаминаза и аспарагиназа, които се намират и в черния дроб:
Asn + H 2 O Asp + NH 3.
Gln + H 2 O Glu + NH 3.
NH3, освободен в тубулите на бъбреците, се неутрализира с образуване на амониеви соли,които се отделят с урината:
NH 3 + H + + Cl - NH 4 Cl.
4 ) Синтез на урея- това е основният начин за неутрализиране на амоняка. Уреята представлява 80% от екскретирания азот.
Процесът на образуване на урея протича в черния дроб и е цикличен процес, наречен " орнитин цикъл"(цикълът на Кребс-Хензелайт).
Цикълът включва две аминокиселини, които не са част от протеините – орнитин и цитрулин, и две протеиногенни аминокиселини – аргинин, аспарагин.
Процесът включва пет реакции: първите две протичат в митохондриите, останалите - в цитозола на хепатоцитите. Някои ензими, образуващи урея, се намират в мозъка, еритроцитите и сърдечния мускул, но целият набор от ензими се намира само в черния дроб.
І реакцияе синтеза на карбамоил фосфат:
CO 2 + NH 3 + 2ATP NH 2 -CO - F + 2ADP + F n.
ензим - карбамоил фосфат синтазаІ (митохондриално-реални). Има и карбамоил фосфат синтаза II (в цитозола), която участва в синтеза на пиримидинови нуклеотиди.
Карбамоил фосфат синтаза I е регулаторен ензим, за който активаторе н– ацетилглутамат.
ІІ реакция– включване на карбамоил фосфат в цикличния процес. При тази реакция той кондензира с орнитин, което води до образуването на цитрулин (реакцията протича и в митохондриите).
IIIреакция- образуване на аргининосукцинат. Това е втората реакция, която използва енергията на АТФ.
IVреакция- разцепване на аргининосукцинат с образуване на аргинин и фумарат. Последният може да влезе в CLC, подобрявайки работата му. Че. това е анаплеротична (попълваща) реакция за CLA.
Vреакция -регенерация на орнитин
С
образуването на урея.
Схема за синтез на урея
CO 2 + NH 3 + 2ATP карбамоил фосфат + 2ADP + F n
1
NH 2 -CO - NH 2
 (урея) орнитин
(урея) орнитин
5 2
аргинин цитрулин
4 3 АТФ
![]() Фумарат AMF
Фумарат AMF
Аргиносукцинат FF n
ензими:
1 - карбамоил фосфат синтаза;
2 - орнитинкарбамоилтрансфераза;
3 - аргининосукцинат синтаза;
4 - аргининосукцинат лиаза;
5 - аргиназа(силни инхибитори на ензима са орнитин и лизин, съперничещи на аргинина, активатори - Ca 2+ и Mn 2+).
Орнитинът, който се възстановява по време на цикъла, може да започне нов цикъл на урея. По своята роля орнитинът е подобен на оксалоацетат в CLA. За преминаването на един цикъл са необходими 3 АТФ, които се използват в 1-ва и 3-та реакция.
Орнитиновият цикъл е тясно свързан с CLA.
Схематично връзката може да бъде представена по следния начин:  2 АТФ
2 АТФ
Орнити- CO 2
нов CLC
цикъл
Фумарат АТФ
аспартат
Това е "двуколесният велосипед" на Кребс - нито едно колело не може да се "върти", без другото да функционира правилно.
Екскрецията на синтезираната урея се осигурява от бъбреците. През деня се отделят 20-35 g урея. Когато количеството протеин в храната се промени, за да се поддържа азотен баланс, скоростта на синтеза на урея в тялото се променя:
протеин с храна синтез на ензими на цикъла синтез на урея,
ако протеинов катаболизъм синтез на урея количество
екскретиран азот.
По време на гладуване и захарен диабет се наблюдава увеличаване на протеиновия катаболизъм и следователно увеличаване на екскрецията на урея.
При заболявания на черния дроб, които са придружени от нарушение на синтеза на урея, концентрацията на амоняк в кръвта се повишава (хиперамонемия) и в резултат на това се развива чернодробна кома.
генетичен дефекти в ензимите за синтез на урея
Известни вродени метаболитни нарушения поради липсата на всеки от петте ензима на цикъла.
При нарушение на синтеза на урея се наблюдава повишаване на концентрацията на амоняк в кръвта - хиперамонемия, която е най-силно изразена с дефект в 1-ви и 2-ри ензими.
Клинични симптоми -общи за всички нарушения на орнитиновия цикъл: повръщане (при деца), отвращение към богати на протеини храни, нарушена координация на движенията, раздразнителност, сънливост, умствена изостаналост. В някои случаи може да настъпи смърт през първите месеци от живота.
Диагнозасе извършват нарушения:
1) чрез определяне на концентрацията на амоняк и междинни продукти от орнитиновия цикъл в кръвта и урината;
2) чрез определяне на активността на ензимите в чернодробни биопсии.
Наследствените ензимопатии на орнитиновия цикъл включват:
хиперамонемияІ Тип – липса на карбамоил-фосфат синтаза I (няколко случая, тежка хиперамонемия);
хиперамонемияІІ Тип – липса на орнитин-карбамоилтрансфераза (много случаи). В кръвта, цереброспиналната течност и урината се повишава концентрацията на амоняк и глутамин.Увеличаването на концентрацията на амоняка води до повишаване на активността на глутамин синтазата;
цитрулинемия– дефект на аргининосукцинат синтазата (рядко заболяване). Голямо количество цитрулин се екскретира с урината, концентрацията на цитрулин в плазмата и цереброспиналната течност се увеличава;
аргининосукцинат ацидурия– дефект на аргинин-сукцинат лиаза (рядко заболяване). Концентрацията на аргининосукцинат в кръвта, цереброспиналната течност и урината се повишава. Заболяването обикновено се развива рано и е фатално в ранна възраст. За диагностициране на това заболяване се използва определянето на наличието на аргининосукцинат в урината (хроматография върху хартия) и еритроцити (по избор). Ранната диагноза се извършва чрез амниоцентеза;
аргининемия - аргиназен дефект. Наблюдава се повишаване на концентрацията на аргинин в кръвта и цереброспиналната течност (еритроцитите имат ниска активност на аргиназа). Ако пациентът бъде прехвърлен на диета с ниско съдържание на протеини, тогава концентрацията на амоняк в кръвта намалява.
Лекция 2
Тема. Специализирани метаболитни пътища
аминокиселини и циклични аминокиселини.
Наследствени ензимопатии
метаболизъм на аминокиселини
Планирайте
1 Метаболитни пътища на безазотния скелет на аминокиселините. Гликогенни и кетогенни аминокиселини.
2 Метаболизъм на глицин и серин.
3 Метаболизъм на аминокиселини, съдържащи сяра. Синтез на креатин.
4 Метаболизъм на аминокиселини с разклонена верига.
5 Метаболизъм на цикличните аминокиселини (фенилаланин, тирозин, триптофан и хистидин).
6 Наследствени нарушения на метаболизма на аминокиселините.
1 Метаболитни пътища на безазотния скелет на аминокиселините. Гликогенни и кетогенни аминокиселини
Скелети на аминокиселини без азот (-кето киселини) се образуват в резултат на реакции на трансаминиране и деаминиране.
Въглеродните скелети на протеиногенните аминокиселини след разцепването на NH2 групата в крайна сметка се превръщат в 5 продукта, които участват в CLA: ацетил-КоА, фумарат, сукцинил-КоА, -кетоглутарат, оксалоацетат.
В CLA въглеродните скелети на аминокиселините са напълно окислени с освобождаването на значително количество енергия, което е съизмеримо с количеството енергия, освободено по време на аеробно окисление 1 молекула глюкоза.
Схематично пътищата за навлизане на α-кето киселини в CLA са показани по-долу:
Ала, Сис, Тре
Ликувай, сър,
PVC
Ацетил-КоА
Ацетоацетил-КоА
Asn, Asp
ОА

Tyr, Fen, Trp
CLC
фумарат
–КИЛОГРАМА
Gln, Glu, Arg, Gis, Pro
Сукцинил-КоА
Ил, Вал, Мет
Гликогенни и кетогенни аминокиселини
Гликогенни аминокиселини- това са аминокиселини, които могат да бъдат субстрати за синтеза на глюкоза, т.к. могат да бъдат превърнати в пируват, оксалоацетат, фосфоенол-пируват - това са съединения прекурсори на глюкоза по време на глюконеогенезата. Тези аминокиселини включват всички протеиногенни аминокиселини с изключение на Lei, Liz.
Кетогенни аминокиселиние субстрат за кетогенезата и синтеза на липиди. Те включват Lei, Liz, Ile, Tir, Trp, Fen. Лей и Лиз са наистина кетогенни аминокиселини, т.к Ile, Trp, Fen могат да бъдат и двете гликогенни.
2 Метаболизъм на глицин и серин
Глицинът се превръща в серин с участието на коензимната форма на фолиевата киселина (BC) - тетрахидрофолиева киселина, или THFA (H4-фолат).
3 Метаболизъм на аминокиселини, съдържащи сяра. Синтез на креатин
Метионине есенциална аминокиселина, която е основният донор на метилови групи в реакциите на метилиране.
Активната форма е S-аденозилметионин (SAM), реакцията на образуване на който е показана по-долу:
Met + ATP S-Аденозилметионин + FFn + Fn.
ензим - метионин аденозилтрансфераза.
SAM участва в реакциите на метилиране по време на синтеза на: холин, креатин, адреналин, меланин, нуклеотиди, растителни алкалоиди. След прехвърлянето на CH 3 групата, SAM се превръща в S-аденозилхомоцистеин, който в резултат на последователност от реакции се редуцира до метионин:
S-аденозилметионин S-аденозилхомоцистеин
аденозин
хранителен метионин
метионинхомоцистеин.
сукцинил-КоА
Този цикличен процес не може да функционира без постоянно снабдяване с Met, т.к Мет се консумира при реакции на катаболизъм.
Мет като донор на метилови групи участва в синтеза на креатин.
Синтез на креатин
Креатинът е основният субстрат за образуването на креатин фосфат в мускулите и нервната тъкан. Синтезът на креатин се извършва последователно в бъбреците и черния дроб (част от него може да се синтезира в панкреаса).
Има два етапа на синтез:
1 Среща се в бъбреците:
Arg + Gln Орнитин + гликоциамин.
(Гуанидиноацетат)
Ензим - глицинамидинотрансфераза (трансаминаза).
2 Появява се в черния дроб след транспортиране на гликоциамин от бъбреците:
S-Аденозилметионин S-Аденозилхомоцистеин
Гликоциамин креатин
ензим - гуанидиноацетат метилтрансфераза.
Освен това, креатинът се фосфорилира с образуването на макроергичен фосфат - креатин фосфат, който е форма на отлагане на енергия в мускулите и нервната система. Ензимът, който катализира тази реакция е креатин фосфокиназа(KFC):
Креатин + ATP Креатин-ph + ADP
неензимно
креатининс урина.
ОНД -Това е неесенциална аминокиселина, чиято основна роля е да:
1) участва в стабилизирането на структурата на протеини и пептиди - образува дисулфидни връзки;
е структурен компонентглутатион трипептид (glu-cis-gli), който като коензим участва във функционирането на антиоксидантната система на организма, транспортирането на определени аминокиселини през мембраните, възстановяването на аскорбинова киселина от дехидроаскорбинова киселина и др.
3) по време на цис катаболизма се образува пируват, който се използва като субстрат за глюконеогенеза, т.е. цис - гликогенна аминокиселина;
участва в синтеза на таурин - физиологично важно съединение, което е необходимо за образуването на сдвоени жлъчни киселини, може да действа като медиатор в централната нервна система и е важно за функционирането на миокарда.
-CO 2
Cys Цистеинова киселина Таурин
CH 2 - CH - COOH CH 2 - CH 2
HO 3 S NH 2 SH NH 2
Тауринът помага за понижаване на нивата на холестерола при атеросклероза, т.к. участва в синтеза на жлъчни киселини.
Аминокиселините с разклонена верига (AKRC) – валин, левцин, изолевцин – се превръщат в -кето киселини (хидрокси киселини с разклонена верига – BCRC) по време на катаболизъм. - NH 3
ACRC OKRC
Етапи на AKRC окисляване:
1) трансаминиране:
AKRC + –KG OCRC + Glu.
ензим - ACRC-аминотрансфераза.
Най-висока активност на този ензим се наблюдава в сърцето, бъбреците, по-малко - в скелетната мускулатура, най-ниската - в черния дроб;
2) дехидратация на SSRC до междинни продукти на CLA. ензим - дехидрогеназа SSRC -локализиран във вътрешната мембрана на митохондриите и катализира реакцията на окислително декарбоксилиране, което води до образуването на CLA междинни продукти:
Leu ацетил-КоА и ацетоацетат.
Вал, Иле сукцинил-КоА.
Катаболизмът на Val и Ile (както и Met) до сукцинил-CoA е придружен от образуването на пропионил-CoA и метилмалонил-CoA:
Метаболизмът като преобразуване на енергия
От молекулярна гледна точка животът на многоклетъчния организъм е сложно организирани генетични и биохимични реакции, които протичат по подреден начин на клетъчно, междуклетъчно, тъканно, органно и системно ниво на организация и отразяват метаболизма или взаимодействието между молекулите и атомите на различни химични съединения (биологични материали).
Реакциите, които протичат по време на метаболизма, се наричат метаболизъмили метаболитни реакции. Както беше споменато в предишната глава, сред основните химични съединения на клетката са: вода (70-90% от обема), която определя свойствата на биологичните материали; соли на Na, Ca, K, Mg, Cl и други микроелементи, разтворени във вода; органични съединения(10-30% от обема), които са ценен вид биологично гориво. Разнообразие от органични молекули се създават чрез комбиниране на въглеродни атоми с атоми на други химични елементи.
Всеки химичен елементхарактеризиращ се с валентност или способност за образуване на определено количество ковалентни връзки. Така се образуват прости молекули:алкохоли (алкохол), включително въглеродна верига и хидроксилна група (HO), амини (аминогрупа - NH 2), киселини (карбоксилова група - COOH) и др. Образуват се различни комбинации от прости молекули сложни молекули,включително молекулите на нуклеинови киселини, протеини, мазнини и въглехидрати, които са най-важни за тялото. Всички тези молекули се образуват в резултат на метаболитни реакции. Известни са два вида метаболитни реакции: анаболенили синтеза на молекули, необходими за живота (анаболизъм) и катаболенили разграждането на молекулите (катаболизъм). Член на метаболитната реакция е метаболит,резултатът от метаболитна реакция е продукт.Ако продуктът от метаболитна реакция служи като изходен материал за следващата реакция, това е така субстрат.
Верига от такива последователни реакции е метаболитен път.Метаболитните пътища са сложни и взаимосвързани. Всякакъв начин
трябва незабавно да се адаптира към текущата ситуация в една клетка и в цялото тяло, въз основа на информация, идваща от други пътища или различни връзки на един път. Всеки метаболитен път се основава на енергийни трансформации, които определят кой път е възможен и кой не.
Енергийни трансформации(нейните трансформации) е една странапривеждане в съответствие на производството на биогорива с постоянно променящите се нужди от биогорива; от друга странатова е подчиняването на скоростта и посоките на енергийна трансформация в отделните клетки на нуждите и жизнения ритъм на целия организъм, който периодично се храни и гладува, работи и почива, носи, храни, расте и учи потомството си да изпитва взаимодействие с околен свят. Освен това в хода на функционирането си всеки организъм, за кратко или продължително, рядко или често, се разболява, изразходвайки запасите от налична енергия през периода на заболяването.
Така енергийните трансформации в клетките, тъканите, органите и системите на тялото се случват постоянно през целия живот на индивида. Условно обаче може да се счита, че те започват с хранене, когато молекулите на хранителните вещества се „разглобяват“ от клетките, за да се получи енергия (биологично гориво), която след това се използва за метаболизъм и синтез на вещества, необходими за организма.
Координацията и интегрирането на всички енергийни трансформации се осигурява от основните регулаторни системи на тялото: нервната и ендокринната, както и имунната система, която проявява регулаторното си действие или индиректно чрез невроендокринна регулация, или чрез лимфоидните си органи, които имат ендокринна функция (вж. Глава 14). Като цяло, контролът на енергийните трансформации в тялото се осигурява от комбинираното действие на невротрансмитери, хормони, растежни регулаторни фактори, както и различни сигнални молекули, които медиират енергийния метаболизъм (вж. Глава 8).
Говорейки за ролята на отделните клетки и тъкани в разпределението и потреблението на енергията, доставяна на тялото под формата на храна, както и за ролята на водата (вж. Глава 6), трябва да подчертаем най-енергийно интензивните и енергийно- консумират клетки на черния дроб, мускулите, мозъка, мастните клетки и червените кръвни клетки. Например при нормално хранене чернодробните клетки съхраняват глюкоза под формата на гликоген, а по време на гладуване я освобождават до изчерпване на целия запас. Ако наличност
гликогенът е изсъхнал, черният дроб превръща аминокиселините в глюкоза (глюконеогенеза), а мазнините (мастните киселини) първо се превръщат в кетонни тела, а след това (чрез окисляване) от тях синтезират триглицериди, които са енергийни носители. Триглицеридите навлизат в кръвния поток и се пренасят до всички тъкани и органи, включително до мозъка, който няма собствени енергийни резерви. Следователно транспортирането на глюкоза в невроните е пасивно и не изисква външно въвеждане на енергия. В пасивната транспортна система участват регулаторни пори, канали на междуклетъчни връзки и клетъчни мембрани (виж глава 6). Тези канали контролират преминаването на различни молекули и йонни потоци, като капацитетът им (например за йони Ca2+, K+ и Na+) зависи от външни за клетката сигнали, пристигащи до портите на канала, в които се намират рецепторите, които ги разпознават.
От своя страна, транспортът на глюкоза в еритроцитите е активен, тъй като се осъществява поради градиент на концентрация или улеснена дифузия поради двупосочното обратно движение на Cl - и HCO - аниони през еритроцитната плазмена мембрана. При което активен транспортвъзниква поради външен източник на енергия, освободен по време на хидролизата на АТФ, и отива (поради това движение) срещу градиента на концентрацията.
Разнообразни източници на енергия за мускулите са глюкоза и гликоген, мастни киселини, кетонни тела и аминокиселини. Специално място във вътреклетъчния енергиен метаболизъм заемат митохондриите, отговорни за "тъканното дишане" или енергийния метаболизъм поради процесите на окисление и фосфорилиране, както и синтеза на АТФ за "критично зависими" тъкани и органи, чието функциониране е напълно зависи от своевременното попълване на резервите от АТФ. Сред такива „критично зависими“ морфофункционални структури са клетките и тъканите на мозъка, миокарда, скелетната мускулатура, ретината, островчетата на Лангерханс в панкреаса и др. Настъпващите в тях нарушения на енергийния метаболизъм имат значителен принос за спектъра и обема на човешките наследствени патология.
В същото време самите митохондрии често „страдат“. И така, към днешна дата е идентифициран голям клас митохондриални заболявания (над 200 нозологии), проявяващи се с увреждане на пациентите поради тежки невродегенеративни симптоми на фона на значително намаляване на енергийните резерви (виж глава 26).
По-специално, нарушения в структурата и функцията на митохондриите са идентифицирани при болести на Алцхаймер и Паркинсон (вж. Глава 28), кардиомиопатичен синдром, захарен диабет и други патологии.
Трофично снабдяване
Трофично снабдяванеили трофизъм, е набор от метаболитни реакции, които определят запазването на структурата и функционирането на клетките, тъканите, органите и системите на тялото, увеличаването им в процеса на прекомерно функционално натоварване (състояние на хипертрофия) или намаляване на процеса на функционално бездействие (състояние на хипотрофия). Трофичното осигуряване на всички телесни структури се осъществява с помощта на нервни влакна и сигнални молекули, които достигат до рецепторите на клетъчните мембрани в целевите клетки. Последните не само информират невроните за тяхното състояние, но и имат стимулиращ ефект върху тях, като инициират адекватни функционални промени в невроните и следователно медиират обратния ефект върху целевите клетки. В същото време тя нервна системаосигурява свой собствен трофизъм единствено благодарение на сигналните молекули, които влизат в невроните от целевите клетки, които те инервират. Всички метаболитни реакции, протичащи в многоклетъчния организъм, се катализират от регулаторни ензимни протеини и протичат поради свързването на ензимите със субстратите. Тези видове реакции се наричат ензимни.
Ензими и ензимни реакции
Ензимната (ензимна) молекула е способна да образува активен център или "джоб", в който влиза субстратната молекула и в който е "атакувана" от различни функционално активни групи.
Според законите на класическата генетика една биохимична реакция се катализира от един ензим. Тя се основава на формулата, предложена през 1941 г. от J. Beadle и E. Tatham: "Един ген - един ензим", която по-късно е трансформирана във формулата: "Един ген - една полипептидна верига", която дълго време се счита за централната догма на молекулярната биология (вж. глава 1).
Трябва да се отбележи, че този принцип, общ за ензимологията, сега доста често се наблюдава за многофункционални ензими и мултиензимни системи (комплекси).
Ензимните реакции, протичащи в тялото, се основават на модели на Michaelis-Menten,който взема предвид всички известни формули за генна експресия:
където Е е ензим; S - субстрат; ES и EP са комплекси на ензима със субстрата S и ензима с продукта P.
По този начин, една ензимна реакция изисква поне субстрат и ензим. В този случай субстратът може да бъде ДНК, РНК, протеин или други молекули, а ензимът може да бъде молекули от регулаторни протеини. Ензимните реакции съпътстват всички алтернативни процеси в клетката и организма: прогрес и регресия, синтез и разпад, развитие и инволюция (стареене), възбуждане и инхибиране, сън и будност и изобщо всички други молекулярни процеси (физикохимични, генетични и биохимични, морфологични , физиологични и патофизиологични), свързани с онтогенезата (вж. Глава 12).
Функциите на клетките и тялото се осигуряват от ензими
Както знаете, ензимите осигуряват множество функции на клетката и тялото. Изброяваме най-важните от ензимните реакции. То:
Експресия на гени, произвеждащи структурни и функционални протеини за клетки, тъкани, органи и телесни системи (реакции на ДНК-тРНК-протеин);
Защитни реакции на организма: вроден и придобит имунитет, коагулация на кръвта, действието на цитохром Р 450 и др. (ензимно-протеинови реакции, ензим-субстрат);
Разпознаване на молекули при трансмембранния им трансфер (включително контрол на йонните потоци), взаимодействие на хормони и други сигнални молекули с рецептори, генериране и провеждане на нервни импулси и др. (същите реакции);
Умствена дейност, работата на мускулните клетки и др. (същите реакции). Освен това това са и ензимни реакции, които протичат между нуклеиновите киселини, започвайки от етапа на оплождане на гаметите. Например, ДНК-ДНК реакцията включва ензими в цитоплазмата на яйцеклетката и ядрени транскрипционни фактори, съдържащи се в яйцеклетката и спермата. Други примери са ДНК-тРНК реакции по време на транскрипция, тРНК-рРНК реакции, тРНК-тРНК реакции по време на транслация и реакции
сайт-специфично свързване между аминокиселинни остатъци на ензими и ДНК нуклеотидни последователности.
ОСНОВНИ вътреклетъчни събития
МЕТАБОЛИЗЪМ
Сред многото събития на вътреклетъчния метаболизъм с различно биологично значение на първо място е необходимо да се разгледат основните "технологични" процеси, протичащи на молекулярно ниво - това е метаболизмът на пуринови и пиримидинови нуклеотиди, несъществени и незаменими аминокиселини, разграждане на ДНК и протеини.
Метаболизъм на нуклеотиди
Нуклеотидът е комбинация от три елемента: фосфат-захар-база. В това съединение фосфатът е свързан да образува енергия. В този случай основният енергиен субстрат, АТФ, играе централна роля, въпреки че участват и други енергийни съединения (GTP, CTP, UTP). Въглехидратните компоненти на нуклеотида са дезоксирибоза или рибоза. Азотните бази са свързани с тях чрез азотния атом (N) на девета позиция на пурина и първа позиция на пиримидина, които осигуряват предаването на наследствена информация.
Азотните основи на нуклеотида са пурини A и G и пиримидини C, T и U.
ДНК нуклеотидите (дезоксирибонуклеотиди) включват: аденин (dAMP), гуанин (dGMP), цитозин (dCMP) и тимин (dTMP).
РНК нуклеотидите (рибонуклеотидите) включват: аденилова (AMP), гуанилова (GMP), цидилова (CMP) и уридилова (UMP) киселини.
Синтез на пуринови нуклеотиди
Повечето клетки синтезират пурини de novo от прекурсори с ниско молекулно тегло. Източници на свободни пурини са нуклеинови киселини, които се разлагат в лизозомите на клетката при разграждането на ДНК молекулата, а мястото на синтеза им е черният дроб.
Пуринов цикъл- това е реакция на риботилиране или добавяне на компонентни фрагменти към рибоза-5-фосфат. Това
механизмът е характерен за синтеза на de novo пурини, свободни пурини и пиримидинови нуклеотиди с участието на 5-фосфорибозил-1-пируват (активната форма на рибоза-5-фосфат) или фосфорибозил-1-пирофосфат (FRPP). Образуването на FRPP става чрез пентозофосфатния път, когато пирофосфатната група (PP) се прехвърля от глутамин ATP, катализиран от FRPP синтетаза. Продуктът от тази реакция е 5-фосфорибозиламин. След образуването му възниква поредица от последователни реакции (общо 9 от тях), завършващи със сглобяването на първия пуринов нуклеотид, включително хипоксантина, инозинова киселина(IMP).
Образуването на IMP служи като своеобразен метаболитен кръстопът: от тази киселина се образува аденин или гуанин.
В хода на тези трансформации от NH 10 -формилтетрахидрофолат или тетрахидрофолат (FH 4) също се образува формилова група (FH) - това е реакция на формилиране. Участник в реакцията на формилиране - тетрахидрофолат (FH 4) е коензим на витамин F или фолиева (птероилглутаминова) киселина. FH 4 се извлича от хидрофолат редуктаза (FH 2) с участието на NADPH и формил трансфераза.
Формилната група също идва от серин, който в присъствието на серин хидроксиметилаза прехвърля хидроксиметилова група (CH2OH) към FH4 и в резултат се образуват глицин, вода и N5N10-метилен-FH4. Последното съединение обаче все още не е готово да участва във формилирането, тъй като метиленовата група (CH2) е по-редуцирана от формиловата група (FH). Следователно CH 2 се окислява от ензима NADP + до метилово производно, което след хидролиза се превръща в N 10 -формил-FH 4 или донор на формилна група, необходим за синтеза на пуринови нуклеотиди. Тук е уместно да се отбележи, че de novo синтезът на пурини не произвежда свободни пурини, тъй като новите пурини незабавно се превръщат в пуринови нуклеотиди. Известен е и път за синтеза на пуринови нуклеотиди, по време на който свободните пурини се превръщат в тях, получени при разпадането на нуклеотидите и запазени след взаимодействие с FRPP. В този път участват две фосфорибозилтрансферази: едната катализира образуването на пуринови нуклеотиди от аденин, а другата от хипоксантин и гуанин. Освен това във втория случай (синтеза на пуринови нуклеотиди от хипоксантин или AMP рибонуклеотид и синтеза на гуанин от GMP рибонуклеотид) участва хипоксантин-гуанин фосфорибозилтрансфераза
(GGPRT), който, взаимодействайки с FRPP, образува IMP и неорганичен фосфор (P). За човешките клетки запазването на аденина изглежда е по-малко важно от запазването на хипоксантина и гуанина. Свободният хипоксантин се образува от AMP, когато фосфатната група (PH) се отстранява от 5-нуклеотидаза и се превръща в аденозин, от който NH 2 групата се отстранява от ензима аденозин деаминаза (ADA). В резултат на това аденозинът се превръща в инозин, от който с помощта на друг ензим се образуват нуклеозид фосфорилаза, хипоксантин и рибоза-1-фосфат.
Трябва също да се отбележи, че при липса на ADA в този метаболитен път се развива автозомно рецесивно лимфоцитно заболяване (20q13.11), проявяващо се с тежък комбиниран имунодефицит (SCID). Предложен в Съединените щати през 1990 г., метод за лечение на SCID е първият случай на използване на метод за генна терапия в медицината, чиято същност е да се въвеждат в костния мозък стволови клетки инвитронормален ADA ген и последваща автотрансплантация на тези клетки in vivo(виж глава 20).
В заключение трябва да се подчертае, че синтезът на пуринови нуклеотиди изисква големи енергийни разходи от клетката и следователно механизмът на повторно използване на свободните пурини е по-полезен за нея, тъй като позволява на клетката да ограничи синтеза de novo.
Освен това в тялото има уникални клетки (еритроцити), които не са в състояние да синтезират пурини de novo и затова използват само готови пуринови бази. Значението на механизма за повторно използване на свободни пурини може да бъде демонстрирано чрез X-свързания рецесивен синдром на Lesch-Nyan (Xq26-27), свързан с умствена изостаналост, нарушена координация и автоагресия (поради липсата на HGPRT). При такива пациенти синтезът de novo на пуринови нуклеозиди в чернодробните клетки е рязко повишен (има захар и основа, но няма фосфат), което води до повишаване на нивото на FRPP, образуване на голямо количество пикочна киселина (в черния дроб) и отлагането на уратни кристали (в бъбреците).
Подобни симптоми се наблюдават и при подагра, но в този случай пациентите нямат неврологични разстройства (по неизвестни причини). Очевидно подаграта е фенокопия на синдрома на Lesch-Nyan. Важно е да се отбележи, че пикочната киселина, която се образува в черния дроб от хипоксантин и гуанин, се инхибира от алопурин. Това лекарство се използва за лечение на подагра, причинява преобладаващо натрупване на гуанин, а не на урат.
и хипоксантин, които са водоразтворими и поради това лесно се отделят от тялото.
Пътят на синтеза на пуринови нуклеотиди de novo е пример за инхибиране на алостерична обратна връзка. В този случай мястото на контрол на инхибирането е първата (обратима) реакция на синтеза на пурин. Той се катализира от FRPP синтетазата, която се инхибира от AMP, ADP, GMP и GDP рибонуклеотиди.
Синтез на пиримидинови нуклеотиди
Повечето клетки синтезират пиримидинови нуклеотиди de novo. В същото време е известен пътят на повторно използване на свободните пиримидини, който е по-малко ярък от този на пурините.
Синтезът на свободни пиримидини започва с аспарагинова киселина и води до образуването на оротова киселина (съединение с циклична структура), която в присъствието на FRPP и под действието на кинази се превръща в урацил (UMP).
В хода на балансирано производство на дезоксинуклеотидни трифосфати (както и пурини) се получава регулиране на алостеричната обратна връзка (виж глава 8). Възстановяването им е на ниво дифосфати с помощта на NADPH (прехвърля електрони към редуктаза). Едва след това те се превръщат в трифосфати на няколко етапа (в резултат на фосфорилиране от кинази с участието на АТФ). Първо, dUTP, който не участва в синтеза (съдържа тимин), се хидролизира в dUMP с образуването на PP. След това dUMP се метилира до dTMP.
Метилиранее прехвърлянето на високоактивна метилова група (CH3) от донор на метионин към молекули на други съединения, включително ДНК (виж по-долу). В този случай нуклеозид-тимидинът се ресинтезира до ТМР чрез действието на тимидин киназа.
Метилирането на dUMP се осъществява и под действието на ензима тимидилат синтетаза, чийто коензим е N 5 N 10 -метилен-FH 4 или метилентетрахидрофолат (FH 4). Освен това, dTMP се фосфорилира до dTTP (метилиран урацил), който се превръща в dTTP.
В случай на синтез на пурин, метиленовата група (CH 2) се окислява, за да образува формилова група (виж по-горе), а в случай на синтез на тимидилат тя се редуцира и прехвърля към метиловата група (CH 3) на тимина в наличието на тимидилат синтетаза. Когато този FH 4 се превърне в FH 2 . Реакцията на обратна редукция изисква ензима дехидрофолат редуктаза, чийто дефицит е свързан с появата на едно от генкопията на фенилкетонурия (виж по-долу).
Друг метаболитен път е превръщането на FH 4 в метилен-FH 4 в реакция със серин, но за това FH 2 се редуцира до FH 4 чрез действието на дехидрофолат редуктаза.
Доказано е, че антифолатите или структурните аналози на фолата, които инхибират FH 2-редуктазата, са антилевкемични лекарства: аметоптерин и аминоптерин, които потискат образуването на dTMP, което е от съществено значение за левкемичните клетки. Освен това образуването на dTMP върви по пътя на инхибиране на FH 2 редуктазата и инхибиране на процеса на превръщане на FH 2 в FH 4 . Освен това, ензимът серин хидроксиметилаза насърчава образуването на метилен-FH (от фолиева киселина), а след това в присъствието на dUMP се образува dTMP.
В допълнение към тези реакции е известна реакция, при която N 5 N 10 - метилен-FH 4 се редуцира до N 5 -метилен-FH 4 , което доставя метилови групи за превръщането на хомоцистеин в метионин в присъствието на метионин синтетаза.
Метионин синтетазата се нуждае от кофактор - витамин B 12, при пълно отсъствие на който се развива автозомно рецесивна пернициозна анемия (6p12-p21.2). При това заболяване тялото не образува стомашния гликопротеин, необходим за възстановяването на витамин B 12 в червата, въпреки че голяма част от него идва от храната. В тази връзка резервите на FH 4 стават недостъпни за синтеза на пурини и тетрахидрофолатът се превръща в метил-FH 4, причинявайки неврологични разстройства при пациенти, свързани с метилмалонова ацидоза. При дефицит на витамин В 12 се развива автозомно рецесивна вродена малабсорбция на фолиева киселина (мегалобластна анемия). Един от неговите кандидат-гени е картографиран към 11q13.3-q14.1.
Известно е, че две реакции зависят от витамин В12. Те се катализират от различни ензими: метилмалонил-КоА-мутаза (6p12-p21.2) и метионин синтаза (генът не е картографиран). При липса на първия ензим се развива фатална ацидоза, докато липсата на втория ензим причинява само ранно забавяне на психомоторното развитие, придружено от неврологични симптоми поради токсичния ефект на хомоцистеина.
Метилиране на ДНК последователности
Метилирането на ДНК последователности (например цитозинови остатъци в позиция 5) протича с образуването на 5-метилцитозин (5-mC) под действието на редица ензими, общо наречени цитозин-ДНК метилтрансферази или М-таза.
М-тазе „поддържащ“ ензим, който разпознава и метилира само полуметилирани ДНК последователности, които се образуват по време на репликация, когато новосинтезираната дъщерна верига все още не е метилирана. Известни са четири такива ензима (Dnmt 1, Dnmt 2, Dnmt 3a и Dnmt 3b). Най-изследваният Dnmt 1 или протеин с молекулно теглооколо 190 kDa, с 2 домейна: каталитичен(разположен на С-края на ензима), структурно подобен на бактериалните цитозинови М-тази, и регулаторни(разположен в N-терминалната част), съдържащ сигнална последователност, която насочва ензима към активни репликационни комплекси в делящите се клетки.
Ензимната активност на Dnmt 1 рязко нараства с началото на синтеза на ДНК. Възможно е генният промотор на този ензим да се активира от продукта на H-ras гена, участващ в трансфера на митогенния сигнал.
Доказано е, че цитозиновите остатъци са основно метилирани като част от CpG динуклеотиди или CpG острови (вж. Глави 1 и 25). Общо около 70% от CpG островите и 6-7% от цитозиновите остатъци са метилирани в еукариотния геном. Това поддържащо метилиране е показано на фиг. 32: В резултат на репликация, метилирани CpG динуклеотиди присъстват в майчината ДНК верига. ДНК метилтрансфераза разпознава метилирани CpGs в нея и пресъздава същия модел на метилиране в дъщерната верига. Трябва да се отбележи, че М-тазите имат само ограничена способност да метилират de novo ДНК последователности в напълно неметилирани региони и да метилират несъответстващи олигонуклеотиди (вж. Глава 10).
В момента са клонирани гени, чиито продукти се проявяват висока способностдо de novo метилиране на ДНК и може да бъде отговорен за този процес.
Метилирането на цитозиновите остатъци засяга структурните характеристики на ДНК, което се проявява в улесняване на прехода на нейните метилирани области от В-форма към Z-форма, увеличаване на стъпката на ДНК спиралата и промяна в кинетиката на образуването на кръстовидни структури. В същото време метиловата група 5-mC изпъква на повърхността на главния жлеб на ДНК във В-формата, което повишава нейната хидрофобност и в някои случаи се превръща в решаващ фактор при взаимодействието на ензимите със съответните области на молекулата на ДНК. Освен това е показано метилиране на други ДНК последователности, като CpNpG, както и
 Ориз. 32.Поддържащо метилиране в генома (съгласно: Herman et al., 1999; http//www.kletca.ru/stem-cells/glossary/)
Ориз. 32.Поддържащо метилиране в генома (съгласно: Herman et al., 1999; http//www.kletca.ru/stem-cells/glossary/)
Сан механизмът на метилиране на аденин и гуанин е метилиране с помощта на сулфониевия катион S-аденизилметионин или SAM. По-специално, механизмът на метилиране на гуанин на позиция 7 (N 7-метилгуанинова група), както и на позиция 2 (OH група на втория, а понякога и третия нуклеотид) играе важна роля за затваряне на тРНК с РНК полимераза II или модифициране на иРНК от 5 "- На това място първият нуклеотид съдържа трифосфатна група и неговият терминален фосфат се отстранява и се заменя с GMP остатък. Благодарение на това затворената иРНК се завършва до функционално активна иРНК. В допълнение към метилирането на аденина и гуанин, за SAM е показано метилиране на аминокиселини и други вещества: креатин, PC и епинефрин (отнася се за катехоламини).
разграждане на ДНК
Разграждането на ДНК е универсален заместител на повечето клетки по време на репликацията на стари молекули за нови. Процесът на разграждане на ДНК се счита за необратим терминален стадий на апоптоза, който се контролира от протеини от семейство В. С 1-2 (виж глава 11).
разграждане на иРНК
Има 2 механизма на разграждане на иРНК (процеси NMD и SMD), свързани с механизмите за възстановяване на клетките (вижте Глава 10).
На фиг. 33 е диаграма на механизма - NMD при бозайници: процесът NMD протича по време на транслация и с негова помощ се съдържа иРНК, съдържаща преждевременни стоп кодони(PSK), който прекъсва транслацията на разстояние от 50-55 нуклеотида напред в хода на четене на последователността екзон-екзон, получена от сплайсинг.
Предшественикът на иРНК (pre-mRNA) в ядрото е свързан с хетеродимера CBP80-CBP20 на основния протеин, свързващ ядрената шапка (CBP).
След образуването на 3" края, пре-тРНК се свързва с ядрения поли (А)-свързващ Upf3a протеин или PABP? протеин (PABP). След това пре-мРНК се снажда и се превръща в иРНК,
 Ориз. 33Схема на NMD при бозайници (по Maquat L., 2005)
Ориз. 33Схема на NMD при бозайници (по Maquat L., 2005)
който се свързва с комплекс, включващ CBP80-CBP20, PABPN1 и цитоплазмения протеин PABPC, и след това се присъединява към протеините на комплекса екзон-екзон или EJC, разположен 20-24 нуклеотида напред. Редица протеини служат като компоненти на EJC, включително:
Пре-мРНК сплайсинг протеини (Pnn/DRS, RNPS1, SRm160, UAP56);
Протеини, участващи в износа на иРНК (REF/Aly, Y14, Magoh);
Протеини, чиито функции не са напълно разбрани (PYM, eIF4AIII и Barentsz/MLN512).
Допълнителни протеини могат също да се прикрепят към EJC комплекса:
NMD фактори (Upf3 или Upf3a, Upf3X или Upf3b, Upf2);
Протеин Upf1 (прикачен, очевидно, временно).
Смята се, че Upf3/Upf3X протеините, които имат предимно ядрена локализация, са в състояние да се движат в цитоплазмата и да взаимодействат с протеина Upf2, който е концентриран по цитоплазмения ръб на ядрената обвивка. Когато се движат, те се образуват първичен комплекс за иницииране на транслацияили mRNP. Този комплекс преминава през първичния цикъл на транслация или във връзка с ядрото или с цитоплазмата, тъй като иРНК служи като техен асоцииран субстрат за NMD.
Процесът на NMD се случва след PSC разпознаване по време на първия цикъл на транслация. Ако транслацията е прекъсната при PSC, който е на повече от 50-55 нуклеотида нагоре от кръстовището екзон-екзон, тогава Upf1 инициира NMD процеса чрез взаимодействие с EJC-асоциирания Upf2 протеин.
Директното разграждане на безсмислени транскрипти в клетки на бозайници се случва както в посоките 5" - 3" и 3" - 5", включително, съответно, декапиране и действието на 5" - 3" екзонуклеотични фактори или деденилиране и действието на 3" - 5" екзозомни фактори.
Участието на EJC комплекси в процеса на NMD се подкрепя от данните, че иРНК, съдържащи PSCs и иРНК, получени от гени без интрони, не се подлагат на NMD. Само новосинтезирани иРНК служат като мишени за NMD, докато стабилните тРНК не се разграждат.
Доказано е, че NMD нарушава редица безсмислени преписи, включително:
иРНК, свързана с алтернативни продукти за сплайсинг;
иРНК, необходима за селенопротеини;
иРНК с отворена рамка за четене;
дали NMD в комплекс с протеин-транслиращи рибозоми или цитоплазмени екстрарибозомни места на разграждане на тРНК.
Ефективността на NMD, като правило, не зависи от локализацията на PSCs и увеличаването на броя на EJC комплексите. Въпреки това, ефективността на NMD може да бъде подобрена чрез други механизми, като моделиране (замяна) на различни генни последователности.
Ролята на NMD в други клетъчни процеси продължава да се изучава. Например, един от неговите фактори - SMG1 - участва в разпознаването и/или възстановяването на увреждането на ДНК, друг фактор - Upfl - участва в безсмислено медиирано алтернативно сплайсинг, както и в наскоро открит нов път за разграждане на иРНК - така нареченото медиирано разграждане на иРНК или SMD. В случай на механизма за разграждане на SMD, РНК-свързващият протеин взаимодейства директно с протеина Upfl, причинявайки разграждане на иРНК на достатъчно отдалечено разстояние от PSC, включително нормалния стоп кодон.
В същото време функционалното предназначение на тези фактори не е окончателно определено за SMD механизма.
Метаболизмът на аминокиселините и неговите нарушения
Аминокиселините се образуват от кето киселини и амоняк (аминогрупи). Те се използват за синтезиране на различни протеини, включително компоненти на клетъчните мембрани, невротрансмитери (напр. 5-хидрокситриптамин и гама-аминобутират или GABA), хормони (например тироксин), хем и други вещества. Източници на аминокиселини са храни и продукти на клетъчния метаболизъм.
Основните аминокиселини са разделени на:
Незаменими (идва изключително отвън), има 10 броя;
Сменяеми (идат отвън и се синтезират в тялото), има и 10 от тях.
Само една аминокиселина - аргинин - е необходима на тялото през ограничен период на развитие (изключително по време на растежа), докато останалите 19 аминокиселини са необходими винаги.
Лизин, фенилаланин и триптофан се считат за абсолютно необходими. Цистеинът може да се получи от фенилаланин, а тирозин от метионин. Незаменимите аминокиселини са или кетогенни (образуващи ацетил-КоА, който се превръща в кетонни тела), или гликогенни (повишаващи нивата на кръвната захар и при диабетици, урината). Въпреки това, левцинът и лизинът са както кетогенни, така и гликогенни.
На фиг. 34 се дава обща схемаметаболизъм на аминокиселини. Както е показано на диаграмата, излишъкът от аминокиселини се образува за сметка на аминокиселинни остатъци, които не се използват в биосинтеза на протеини или за други нужди на клетката.
Полученият излишък от аминокиселини под формата на метаболитен фонд се използва (при разграждането им) за производство на енергия и създаване на енергийни резерви (мазнини и гликоген), а аминният азот се отделя с урината под формата на урея.
Ако е необходимо, функционалните мускулни протеини могат да се превърнат в резерви за производството на аминокиселини (има повечето от тях в тялото). Редица наследствени заболявания, свързани с нарушения в метаболизма на аминокиселините, са идентифицирани при хората (вж. Глава 21). За пациенти с такива нарушения е характерен или дефицит, или излишък на определена аминокиселина, което води до
 Ориз. 34.Обща схема на метаболизма на аминокиселините (според Elliot V., Elliot D., 2002)
Ориз. 34.Обща схема на метаболизма на аминокиселините (според Elliot V., Elliot D., 2002)
лошо храносмилане и усвояване на храната, изчерпване на организма, забавено психомоторно и физическо развитие, оток на тъканите, неврологични и други симптоми, базирани на синтеза на дефектни протеини.
Повечето типичен примерТези заболявания са автозомно рецесивна фенилкетонурия (PKU), която се развива в резултат на дефицит на ензима фенилаланин-4-хидроксилаза
PKU има редица генокопи (най-често злокачествени), които се развиват в резултат на дефицит на 6-пирувоилтетрахидроптерин синтаза (11q22.3-q23.3), цитозолна дихидроптеридин редуктаза (4p15.31), цитозолна гуанозин циклохидролаза (нециклохидролаза) на цитозола , или тетрахидробиоптерин кофактор BH 4 хидролази на ароматни аминокиселини: фенилаланин, тирозин и триптофан (генът също не е картографиран).
Това също включва генокопие на PKU поради дефицит на дехидрофолат редуктаза.
Преди да разгледаме механизмите на патогенезата на генокопията на фенилкетонурия, отбелязваме, че в нормално функциониращ организъм фенилаланинът не се подлага на деаминиране, а се превръща в тирозин (под действието на фенилаланин-4-хидроксилаза).
При PKU синтезът на тирозин е възпрепятстван или напълно блокиран и фенилаланинът е „принуден“ да се деаминира до фенилпируват (кето киселина), който се екскретира с урината.
Във връзка с тези особености на метаболизма на фенилаланина в организма и наличието на генокопи на PKU, беше направен анализ на състава на свободните аминокиселини и резултатите от метаболизма на фенилаланина в кръвната плазма на небременни и бременни жени. - носители на гена PKU (Vasilyeva O.V., 1999). Съставът на свободните аминокиселини в кръвния серум на небременни жени (първа група)се характеризира със съотношението на есенциални и неесенциални аминокиселини съответно 38 и 62%, както и съотношението на хидрофобни и неутрални към други аминокиселини, възлизащо на 71 до 29%. Отбелязани са най-високи концентрации на аланин, треонин, лизин и аргинин; най-малките - аспарагинова и глутаминова киселини.
При анализ на корелациите между количествените характеристики на аминокиселинния спектър бяха идентифицирани три степени на конюгиране на количествените показатели.
lyable системи: високи, средни и ниски нива на асоцииране на аминокиселини.
респективно високи ниваса лизин, фенилаланин и тирозин; средните нива са хистидин, цистеин и валин; ниските нива са левцин, изолевцин, метионин, аланин и аспарагинова киселина. В допълнение към тези три нива бяха идентифицирани нивата на корелации, характерни за аргинин и глутаминова киселина, както и за серин и треонин. Получените данни бяха обяснени от автора със структурните и функционални характеристики на аминокиселините:
Фенилаланинът е предшественик на тирозин;
Фенилаланин, тирозин и лизин участват в синтеза на ацетил-КоА без междинно образуване на пируват;
Аргинин, хистидин и валин могат да участват в синтеза на глутаминова киселина;
Левцин, изолевцин и метионин са хидрофобни аминокиселини;
Аланинът се образува по време на трансаминирането на пируват, който може да стане основа за синтеза на глицин; аспартатът служи като донор на аминогрупата.
Физиологичната бременност доведе до промени във фонда на свободните аминокиселини поради намаляване на съдържанието на глицин, валин, левцин през първия триместър; през втория триместерегистидин, фенилаланин и цистеин. Ако според масовия (пилотен) скрининг на бременни жени нивото на фенилаланин в кръвната плазма съответства на 1,2 mg%, то това се оценява като критерий за избор на жени в групата на „потенциални хетерозиготи“ за PKU гена .
Ако експресията на PKU гена, скрит в тялото на майката, доведе през първия триместър на бременността до повишаване на нивото на фенилаланин в кръвната плазма над 10 mg%, то това се оценява като причина за нарушения в развитието на плода.
Задължително хетерозиготно носителство на PKU гена при майки, родили деца с PKU (втора група),в сравнение с жени без PKU ген (трета група)проявява се с нарушения в метаболитния фонд от аминокиселини под формата на висока концентрация на глутаминова и аспарагинова киселини, треонин и глицин.
Отбелязвайки голямото теоретично и практическо значение на работата на O.V. Василиева, можем да заключим, че съвременните идеи за спектъра и механизмите на проявление на различни генокопи
PKU (и евентуално генкопия при други наследствени метаболитни заболявания - NBO аминокиселини) трябва да бъдат значително разширени. Това може да стане не само чрез анализиране на фонда от всички аминокиселини, групирани в отделни групи в зависимост от степента на конюгиране с техните структурни и функционални характеристики, но и чрез анализиране на ролята на ензимните протеини, участващи в метаболизма на тези аминокиселини. Това заключение се подкрепя и от пример за автозомно рецесивна левциноза или заболяване на урината от кленов сироп, при което бяха идентифицирани три генокопия поради ензимен дефицит: алфа-кето киселинни дехидрогенази с различни странични вериги, тип I A (19q13.1-13.2), тип I B (6p21-p22) и тип II (1p31). Левцинозата се развива в резултат на нарушение на окислителното декарбоксилиране на алфа-кето киселини, придружаващи образуването на алифатни аминокиселини: левцин, изолевцин и валин.
Завършвайки разглеждането на данните за значението на метаболизма на отделните аминокиселини, важно е да се отбележи, че заедно с дефицит може да се наблюдава и техният излишък и след това пациентите се диагностицират, например, със заболявания като:
алкаптонурия(3q2) - резултат от нарушено разцепване на тирозин поради излишък от хомогентизинова киселина, която е неговият дифенолен продукт; в този случай дифенолът се комбинира с атмосферния кислород и образува пигмент, поради което урината става тъмна;
цистаурия(16q) е излишък от цистатион (виж по-долу).
Синтез на аминокиселини
Прекурсорите на основните аминокиселини са 5 химични съединения: алфа-кетоглутарат, 3-фосфоглицерат, оксалоацетат (R= CH 2 COO), фосфоенол-пируват, пируват (R=CH 3) и два монозахарида на пентозния път на phospha. Помислете за механизмите на тяхното превръщане в аминокиселини.
Аминокиселини като продукти на метилиране
Аминокиселините могат да бъдат продукти на метилиране или прехвърляне на метилова група от донор на метионин към различни съединения (вижте по-горе). Когато метионинът реагира с АТФ, неговата NH3+ група се активира, за да образува сулфониев катион или S-аденизилметионин (SAM). Прехвърлянето на метиловата група се катализира от трансметилази.
По време на тази реакция три фосфатни групи на АТФ се превръщат в пирофосфат (PP) и неорганичен фосфор (P), след което PP се разделя на две P молекули.
Първо, SAM се превръща в S-аденозилхомоцистеин, който се превръща в хомоцистеин-метионин (има SH група или тиолова група, вместо S-CH3 групата). След това тиоловата група се прехвърля от хомоцистеин към серин, за да се образува цистеин. Междинният продукт в тази реакция е цистатионът, чийто излишък се екскретира с урината.
Продуктите за метилиране на SAM са още: креатин, фосфолипид - PC и катехоламин - адреналин.
Аминокиселини като продукти на трансаминиране
Аминокиселините могат да бъдат продукти на трансаминиране или дезаминиране. Например, глутаминовата киселина се синтезира от глутамат дехидрогеназа, чиито кофактори са NAD+ и NADP+. Тази реакция е обратима.
Донори на глутаминова киселина са аспарагинова киселина и аланин, които се образуват при трансаминирането на оксалоацетат и пируват. Глутаминовата киселина се деаминира чрез елиминиране на два водородни атома в присъствието на глутамат дехидрогеназа, като се използва NAD+ или NADP+ като окислител. Този ензим се инхибира алостерично от ATP и GTP (те показват големи запаси на енергия), но се активира от ADP и GDP (те показват липса на енергия).
След дезаминиране на глутаминовата киселина се образува алфа-кетоглутарат, който участва в Цикъл на Кребс(цикъл на лимонена киселина), който позволява окисляването на глутаминовата киселина
до H2O и CO2.
Тъй като алфа-кетоглутаратът се превръща в оксалоацетат, той може да участва в синтеза на глюкоза, т.е. глутаминовата киселина е гликогенна аминокиселина.
За други аминокиселини (с изключение на глутамин) няма съответни дехидрогенази. Следователно тяхното дезаминиране протича не на един, а на два етапа: първият етап е трансаминиране, вторият етап е дезаминиране. По принцип този метаболитен път, общ за всички аминокиселини, се нарича трансаминиранеили дезаминиране.
Трансаминирането се катализира от аминотрансферази (трансаминази), които са специфични за различните аминокиселини.
Активните центрове на трансаминазите съдържат коензима пиридоксал-5-фосфат (РР), който действа като електрофилен междинен продукт, който първо приема аминогрупа
(служи като негов акцептор), а след това (като донор) го прехвърля към кето киселината.
Работната група на PF е алдехидната група (CHO).
PF включва три производни на витамин B 6: пиридоксал, пиридоксин и пиридоксамин.
Механизмът на трансаминиране може да бъде демонстриран с примера на аланин, който е транспортната форма на аминоазот в кръвта.
Около 30% от аминоазотът е концентриран в аланин, който влиза в черния дроб след разграждането на мускулните протеини и се образува от пируват при трансаминирането на други аминокиселини.
Когато аланинът и алфа-кетоглутаратът взаимодействат, се образува пируват, който реагира с глутамат, това е първи етап. Във втория етапГлутаматът добавя NAD+ и вода, за да образува кетоглутарат, който реагира с NaPH+ и NH 4 . Аланинът се деаминира в черния дроб. Полученият амоняк отива за синтеза на урея, а пируватът - за синтеза на глюкоза, която се връща с кръвта в мускулите, затваряйки глюкозо-аланиновия цикъл на пренос на амоняк. Този цикъл е от особено значение по време на гладуване, когато по време на глюконеогенезата в черния дроб се използват аминокиселини от разграждането на мускулните протеини.
Синтез на серин и глицин
Серинът се синтезира на три етапа от гликозил-3-фосфат глицерол, който първо се окислява до кето киселина (трифосфат хидроксипируват); той се трансаминира от глутаминова киселина и се превръща в 3-фосфат-серин, който се хидролизира до серин и неорганичен фосфор.
Глицинът се синтезира чрез отстраняване на хидроксиметиловата група от серина (виж по-горе). Реакцията идвас участието на тетрахидрофолиева киселина, която е носител на моновъглеродни групи.
Този вид трансфер е важен за синтеза на нуклеотиди.
Синтез на други аминокиселини
Глутаминът (както и аланинът) служи като транспортна форма на амоняк в кръвта, който се образува по време на дезаминирането на аминокиселините.
Амонякът е токсичен и следователно навлиза в черния дроб не в свободна форма, а в комбинация с глутаминова киселина, образувайки амид на глутаминова киселина с участието на ензима глутамин синтетаза, или
глутамин. Като междинен продукт гама-глутамил фосфат (анхидрит на глутамин и фосфорна киселина) е високоенергийно съединение, което може да взаимодейства с амониеви йони с участието на синтетаза (виж глава 8).
Източникът на глутамин е алфа-кетоглутарат от цикъла на Кребс, който претърпява трансаминиране с други аминокиселини.
Глутаминът се транспортира чрез кръвта до черния дроб, където се хидролизира от глутаминаза, а амонякът, освободен при този процес, се използва за синтеза на урея.
Фенилаланинът е ароматна аминокиселина, чийто излишък в нормално функциониращ организъм се превръща в тирозин с участието на фенилаланин-4-хидроксилаза, която доставя 2 водородни атома от коензима тетрахидробиоптерин (виж по-горе).
Левцин, изолевцин и валин са алифатни аминокиселини, чиито междинни продукти се натрупват като кето киселини при левциноза (виж по-горе).
Метаболизъм на други съединения от аминокиселини
Освен протеини от аминокиселини се образуват: амини (в резултат на декарбоксилиране); катехоламини или хормони, подобни по структура на катехола (напр. 1,2-дехидроксибензен), включително допамин, епинефрин и норепинефрин; невротрансмитери (GABA и 5-хидрокситриптамин), както и хормона - тироксин.
Аминогрупите след отстраняването им от аминокиселините се екскретират в урината под формата на урея - това е инертно водоразтворимо нетоксично съединение.
Уреята се образува в черния дроб, когато гуанидиновата група се отцепи от аргинин. В този случай по пътя се образува аминокиселина - орнитин, която не е част от основните протеини на тялото.
За да се превърне орнитин обратно в аргинин, се използват въглероден атом, получен от въглероден диоксид и аминоазот, освободен по време на метаболизма на някоя от основните аминокиселини.
Образуването на аргинин от орнитин протича на няколко етапа. Като междинен продукт се образува аминокиселина - цитрулин, която също не е част от основните протеини на тялото; той стимулира синтеза на урея в черния дроб (като орнинтин и аргинин).
Разграждане на протеини
Разграждането на протеина е замяната на старите протеинови молекули с нови молекули. Той се среща във всички клетки и тъкани на тялото по време на метаболизма. Протеините имат различна продължителност на живота. Дългоживеещите протеини са структурни протеинии хемоглобин. Чернодробните протеини живеят няколко дни.
Много протеини имат живот не повече от 20 часа, а някои от тях живеят не повече от десет или дори две минути.
Поради различната продължителност на живота на протеините, тяхното разграждане се характеризира с висока селективност.
По време на синтеза на аминокиселини и последващото производство на структурни и регулаторни протеини от тях не винаги се спазва тяхната абсолютна структурна (и функционална) точност. Следователно в клетките неизбежно се образуват погрешни аминокиселини, което води до неправилно нагъване на протеини (виж глава 3), а такива протеини се разрушават от клетката, т.е. преминават през процес на унищожаване. За селективното унищожаване е важен протеин, убиквитин. Това е малък протеин, участващ в АТФ-зависима реакция, при която неговата крайна карбоксилна група се свързва с аминогрупата на страничната верига на целевия протеин (лизинови остатъци), която трябва да претърпи разграждане, т.е. той сякаш "маркира" за нея.
Изпратете вашата добра работа в базата от знания е лесно. Използвайте формуляра по-долу
Студенти, специализанти, млади учени, които използват базата от знания в своето обучение и работа, ще Ви бъдат много благодарни.
Хоствано на http://www.allbest.ru/
МОСКОВСКИЯ ДЪРЖАВЕН ТЕХНИЧЕСКИ УНИВЕРСИТЕТ на името на Н.Е. БАУМАН
Факултет по биомедицинско инженерство
Катедра Медико-технически информационни технологии
Метаболизмът на аминокиселините и неговата роля в живота на организма
(по биохимия)
Евдокимова М.П. Група: BMT2-32
Ръководител: Йершов Ю.А.
Москва 2012г
Концепция за аминокиселини
Метаболизъм на аминокиселини
Основни пътища на метаболизма на аминокиселините
Деаминиране
Трансдеаминиране
Декарбоксилиране
Нарушаване на метаболизма на аминокиселините
Заключение
метаболизъм на органични съединения аминокиселина тирозин
Цел: Описват пътищата на метаболизма на аминокиселините и определят значението на метаболитния процес.
Концепцията за аминокиселини
Аминокиселините са най-важни, а някои от тях са жизненоважни органични съединения, чиято молекула едновременно съдържа карбоксилни и аминогрупи.
Аминокиселините изпълняват много функции в живите организми. Те са структурни елементи на пептиди и протеини, както и на други природни съединения. Всички протеини, независимо дали са от най-древните линии на бактерии или от по-висши организми, използват един и същ набор от 20 различни аминокиселини, ковалентно обвързан приятелс приятел в определена последователност, характерна само за даден протеин. Наистина забележително свойство на клетките е способността им да комбинират 20 аминокиселини в различни комбинации и последователности, което води до образуването на пептиди и протеини с напълно различни свойства и биологична активност. От едни и същи градивни елементи различните организми са в състояние да произвеждат толкова разнообразни продукти като ензими, хормони, протеин на очната леща, пера, паяжини, млечни протеини, антибиотици, гъбични отрови и много други съединения, надарени със специфична активност. Също така, някои от аминокиселините са невротрансмитери или прекурсори на невротрансмитери, невротрансмитери или хормони.
Метаболизъм на аминокиселини
Най-важната и незаменима роля в живота на организмите играе метаболизмът на аминокиселините. Непротеиногенните аминокиселини се образуват като междинни продукти по време на биосинтеза и разграждане на протеиногенни аминокиселини или в цикъла на уреята. Освен това за животните и хората аминокиселините - градивните елементи на протеиновите молекули - са основни източници на органичен азот, който се използва предимно за синтеза на специфични за тялото протеини и пептиди, а от тях - азотсъдържащи вещества на небелтъчна природа (пуринови и пиримидинови бази, порфирини, хормони и др.).
Ако е необходимо, аминокиселините могат да служат като източник на енергия за тялото, главно поради окисляването на техния въглероден скелет.
Основните направления на метаболизма на аминокиселините.
Привидно постоянство химичен съставна живия организъм се поддържа благодарение на баланса между процесите на синтез и разрушаване на съставните му компоненти, т.е. баланс между катаболизъм и анаболизъм. В растящия организъм този баланс се измества към протеиновия синтез, т.е. анаболната функция преобладава над катаболната. В тялото на възрастен, в резултат на биосинтеза, ежедневно се актуализират до 400 g протеин. Освен това различните протеини се актуализират с различна скорост - от няколко минути до 10 или повече дни, а такъв протеин като колаген практически не се актуализира през целия живот на тялото. Като цяло полуживотът на всички протеини в човешкото тяло е около 80 дни. От тях около една четвърт от протеиногенните аминокиселини (около 100 g) се разграждат необратимо, които трябва да се обновяват за сметка на хранителните протеини, останалите аминокиселини се синтезират частично от организма. При недостатъчен прием на протеини от храната тялото използва протеините на някои тъкани (черен дроб, мускули, плазма и др.) за насочен синтез на протеини на други жизненоважни органи и тъкани: сърдечен мускул и др. Биосинтезата на протеин се извършва само ако всичките 20 естествени аминокиселини присъстват като първоначални мономери, всяка в точното количество. Продължителното отсъствие и недостатъчен прием дори на една от 20-те аминокиселини води до необратими промени в организма.
Протеините и аминокиселините са най-важните азотсъдържащи съединения на животинските организми – те представляват повече от 95% от биогенния азот. Концепцията за азотен баланс (АВ) е неразривно свързана с метаболизма на протеините и аминокиселините, което се разбира като разликата между количеството азот, въведен в тялото с храна (Nin) и количеството азот, отделяно от тялото ( Nout) под формата на крайни продукти на азотния метаболизъм, главно урея:
AB \u003d N вход - N изход, [g ден -1]
При положителен азотен баланс, биосинтезата на протеините преобладава над процесите на техния разпад, т.е. по-малко азот се отделя от тялото, отколкото влиза. Положителен азотен баланс се наблюдава в периода на растеж на тялото, както и по време на възстановяване от изтощителни заболявания. При отрицателен азотен баланс разграждането на протеините надделява над синтеза им и повече азот се отделя от тялото, отколкото влиза. Това състояние е възможно при стареене на организма, гладуване и различни изтощителни заболявания. Обикновено практически здрав възрастен има азотен баланс, т.е. количеството азот, въведен в тялото, е равно на количеството, което се отделя. Нормите на протеин в храната при достигане на азотен баланс са средно 100-120 g·ден -1.
Абсорбцията на свободни аминокиселини в резултат на хидролизата на протеините се извършва главно в тънките черва. Този процес представлява активен транспорт на аминокиселинни молекули, който изисква енергия и зависи от концентрацията на Na+ йони. Открити са повече от пет специфични транспортни системи, всяка от които транспортира най-близките по химическа структура аминокиселини. Различни аминокиселини могат да се конкурират помежду си за местата на свързване върху транспортните протеини, вградени в мембраната (вижте Глава 15 от този раздел). Така абсорбираните аминокиселини в червата влизат в черния дроб през порталната система и след това влизат в кръвта.
По-нататъшният катаболизъм на аминокиселините до крайните продукти е комбинация от реакции на дезаминиране, трансаминиране и декарбоксилиране. В същото време всяка отделна аминокиселина има свой специфичен метаболитен път.
Дезаминиране на аминокиселини
Дезаминирането е отстраняването на аминогрупите от аминокиселините за образуване на амоняк. Именно с реакциите на дезаминиране най-често започва катаболизмът на аминокиселините. В живите организми са възможни четири вида дезаминиране на аминокиселини.
Общият продукт на всичките четири вида дезаминиране е амонякът, съединение, което е доста токсично за клетките и тъканите, така че се детоксикира в тялото (вижте по-долу). В резултат на дезаминиране поради "загубените" аминогрупи под формата на амоняк общият брой на аминокиселините намалява. Повечето живи организми, включително хората, се характеризират с окислително дезаминиране на аминокиселини, докато други видове дезаминиране се срещат само в някои микроорганизми.
Окислителното дезаминиране на L-аминокиселините се осъществява от оксидазите, присъстващи в черния дроб и бъбреците. Често срещан коензим на L-аминокиселинната оксидаза е FMN, който действа като водороден носител от аминокиселина към кислород. Цялостната реакция на окислително деаминиране е както следва:
R-CH(NH2)-COOH + FMN + H2O >
> R-CO-COOH + FMNN 2 + NH 3 + H 2 O 2
По време на реакцията се образува междинно съединение - иминокиселина, която след това се хидратира, за да се образува кето киселина. В допълнение към кетокиселината и амоняка, като основни продукти на деаминирането, в тази реакция се образува и водороден прекис, който след това се разлага на вода и кислород с участието на каталаза:
H 2 O 2 > H 2 O + SO 2
Окислителното дезаминиране, като самостоятелен процес, играе незначителна роля в превръщането на аминогрупите на аминокиселините; само глутаминовата киселина се деаминира с висока скорост. Тази реакция се катализира от ензима глутамат дехидрогеназа, чийто коензим е NAD или NADH. Активността на глутамат дехидрогеназата се регулира от алостерични модификатори, GTP и ATP действат като инхибитори, а GDP и ADP действат като активатори. Окислителното деаминиране на глутаминовата киселина може да бъде представено със следната схема:
HOOS-CH 2 -CH 2 -CH (NH 2) -COOH + NAD>
> HOOS-CH 2 -CH 2 -CO-COOH + NH3 + (NADH + H +)
Тази реакция е обратима, но в условията на жива клетка равновесието на реакцията се измества към образуването на амоняк. Други, неокислителни видове дезаминиране са характерни за серин, цистеин, треонин и хистидин. Останалите аминокиселини се подлагат на трансдеаминиране.
Трансдеаминиране. Трансдеаминирането е основният път за катаболното разграждане на аминокиселините. По името на процеса е лесно да се предположи, че протича на два етапа. Първият е трансаминиране, а вторият е окислителното дезаминиране на самата аминокиселина. Трансаминирането се катализира от ензима аминотрансферази, известен също като трансаминази. Пиридоксал фосфатът (витамин В6) действа като аминотрансферазен коензим. Същността на трансаминирането е прехвърлянето на аминогрупа от b-аминокиселина към b-кето киселина. По този начин, реакцията на трансаминиране е междумолекулен редокс процес, включващ въглеродни атоми не само на взаимодействащите аминокиселини, но и на пиридоксал фосфат.
Декарбоксилиране на аминокиселини
Декарбоксилирането на аминокиселини е процес на отстраняване на карбоксилна група от аминокиселина под формата на CO2. Някои аминокиселини и техните производни могат да претърпят декарбоксилиране в условията на жив организъм. Декарбоксилирането се катализира от специални ензими - декарбоксилази, чийто коензим (с изключение на хистидин декарбоксилазата) е пиридоксал фосфат. Продуктите на декарбоксилиране са амини с биологична активност - биогенни амини. Повечето от невротрансмитерите и регулаторните фактори на локално действие (тъканни медиатори, които регулират метаболизма) принадлежат към тази група съединения. Реакцията на декарбоксилиране на произволна аминокиселина може да бъде представена, както следва:
Декарбоксилаза
Образуване на биологично активни амини
Раздел. предшественици, химическа структура, биологична ролябиогенни амини
Нарушения на метаболизма на аминокиселините
Метаболизмът в тялото е много важен процес. Всяко отклонение от нормата може да доведе до влошаване на човешкото здраве. Има наследствени и придобити нарушения на метаболизма на аминокиселините. Най-висока скорост на метаболизма на аминокиселините се наблюдава в нервната тъкан. Поради тази причина в невропсихиатричната практика различни наследствени аминоацидопатии се считат за една от причините за деменция.
Нарушаване на метаболизма на тирозина.
Тирозинът, освен че участва в синтеза на протеини, е предшественик на надбъбречните хормони адреналин, норепинефрин, допаминов медиатор, хормони на щитовидната жлеза, тироксин трийодтиронин, пигменти. Нарушенията на метаболизма на тирозина са многобройни и се наричат тирозинемии.
Тирозинемия тип I.
Етиология. Заболяването се появява, когато има дефицит фумарилацетоацетат хидролази. В същото време фумарилацетоацетатът и неговите метаболити се натрупват, засягайки черния дроб и бъбреците.
клинична картина.
остра форма представлява по-голямата част от случаите с начало между 2 и 7 месечна възраст. и смърт при 90% от пациентите на възраст 1-2 години поради чернодробна недостатъчност.
В хронична форма болестта се развива по-късно, прогресира по-бавно. Продължителността на живота е около 10 години. Основи на лечението . Лечението е неефективно. Използва се диета с намаляване на количеството протеин, фенилаланин и тирозин, инжекции с глутатион. Необходима е чернодробна трансплантация.
Тирозинемия тип 2. Много по-рядко заболяване.
Етиология. Заболяването възниква, когато има дефицит на тирозин аминотрансфераза.
клинична картина. Умствено и физическо изоставане, микроцефалия, катаракта и кератоза на роговицата (псевдохерпетичен кератит), кожна хиперкератоза, самонараняване, нарушена фина координация на движенията.
Основи на лечението . Диета с ниско съдържание на тирозин е ефективна и лезиите на кожата и роговицата изчезват бързо.
Тирозинемия при новородени.
Етиология. Неонаталната тирозинемия (тип 3) е резултат от дефицит на хидроксифенилпируват хидроксилаза. По-често се наблюдава при недоносени бебета.
клинична картина. Намалена активност и летаргия. Аномалията се счита за безобидна. Дефицитът на аскорбинова киселина засилва клиничната картина.
Основи на лечението. Диета с намаляване на количеството протеин, фенилаланин, тирозин и високи дози аскорбинова киселина.
Алкаптонурия.
Етиология. Генетична автозомно рецесивна ензимопатия. Заболяването се основава на намаляване на активността на чернодробния ензим хомогентизат оксидаза, в резултат на което в тялото се натрупва хомогентизинова киселина.
клинична картина. Тъй като хомогенизаторът полимеризира във въздуха в меланиноподобно съединение, най-честият и постоянен симптом е тъмна урина, тъмнокафяви петна остават по пелената и бельото. В противен случай болестта не се проявява в детска възраст.
С възрастта хомогентизиновата киселина се натрупва в образуванията на съединителната тъкан, склерата и кожата, причинява дълбока сянка на ухото и носния хрущял, оцветяване на областите на дрехите, изпотяване на части на тялото (подмишници).
В същото време хомогентизиновата киселина инхибира лизил хидроксилазата, предотвратявайки синтеза на колаген, което прави хрущялните образувания крехки. В напреднала възраст се развива дегенеративна артроза на гръбначния стълб и големите стави, междупрешленните пространства се стесняват.
Основи на лечението. Макар че ефективни начининеизвестно, по аналогия с други аминокиселинни нарушения, се препоръчва с ранна възрастограничаване на приема на фенилаланин и тирозин, което трябва да предотврати развитието на охроноза и ставни нарушения. Предписвайте големи дози аскорбинова киселина за защита на активността на лизил оксидазата.
албинизъм
Етиология. Заболяването се причинява от пълен или частичен дефект в синтеза на ензима тирозиназа (честота 1:20 000), който е необходим за синтеза на дихидроксифенилаланин в пигментните клетки.
клинична картина. При пълно отсъствие на ензима, тоталната делигментация на кожата, косата, очите и цветът е еднакъв за всички расови групи и не се променя с възрастта. Кожата не потъмнява, невуси, старчески петна напълно липсват, развива се фотодерматит. Силно изразен нистагъм, фотофобия, дневна слепота, червен зеничен рефлекс. При частична недостатъчност се забелязват светложълта коса, леко пигментирани бенки и много светла кожа.
паркинсонизъм .
Етиология. Причината за паркинсонизъм (честота след 60 години 1:200) е ниската активност на тирозин хидроксилазата или DOPA декабоксилазата в нервната тъкан, като същевременно се развива дефицит на невротрансмитера допамин и натрупване на тирамин.
клинична картина. Най-честите симптоми са мускулна скованост, скованост, треперене и спонтанни движения.
Основи на лечението. Необходими са системно приложение на лекарствени аналози на допамина и употребата на инхибитори на моноаминоксидазата.
Фумарат ацетоацетат
Фенилкетонурия
Етиология. дефицит фенилаланин хидроксилаза.Фенилаланинът се превръща във фенилпируват.
клинична картина.
§ Нарушение на миелинизацията на нервите
§ Мозъчната маса е под нормалната.
§ Умствена и физическа изостаналост.
Диагностични критерии:
§ нивото на фенилаланин в кръвта.
§ Тест за FeCl3.
§ ДНК проби (пренатално).
Заключение
Метаболизмът на протеините и аминокиселините играе важна и незаменима роля в живота на организмите. Това е механизъм, усъвършенстван до най-малкия детайл. Изучаването на протеиновия метаболизъм ни позволява да разберем в детайли дълбокия смисъл, присъщ на най-важния биологичен постулат, който казва, че „организмите стават протеини“. Този постулат съдържа изключителното биологично значение, което е присъщо изключително на протеиновите съединения.
Основна литература
1. Ершов Ю.А., Зайцева Н.И. Основи на биохимията за инженери. MSTU 2010 г
2. Ершов Ю. А. и др. обща химия. М. 2011 г.
3. Ленингер А. Основи на биохимията. М. Мир. 1985. 1055 с.
4. Николаев А. Я., Биологична химия, М. "Агенция за медицинска информация", 2004г.
5. Флорентиев В. Л., Биохимия. - М., 2004. - 464 с.
6. Березов Т.Т., Коровкин Б.Ф., Биологична химия. М, Медицина, 1998
7. Ершов Ю.А. и др. Обща химия. 8-мо изд. М. VS. 2009. 560 стр.
8. Ершов Ю.А. и др. Кинетика и термодинамика на биохимичните и физиологични процеси. М. Медицина. 1990. 208 с.
9. Колман Я., Рем К.-Г. Визуална биохимия. М., Мир, 2004. 269 с.
Хоствано на Allbest.ru
...Подобни документи
Процесът на метаболизъм на протеини, аминокиселини и отделни аминокиселини. Биогенни амини, тяхната роля и значение. Окисление на биогенни амини (моноамин оксидаза). Ролята на хистамина в развитието на възпаление и алергични реакции. Антихистамини, техните задачи и функции.
презентация, добавена на 13.04.2015
Изучаване на хормоните - производни на аминокиселини, особеностите на техния синтез и механизма на действие на клетката. Физиологичната роля на катехоламините и тяхната функция - мобилизиране на защитните сили на организма при стрес. Анализ на ефекта върху секрецията.
тест, добавен на 27.02.2010 г
Ролята на минералите за осигуряване на нормалното протичане на жизнените процеси на човешкото тяло. Препарати, съдържащи макро- и микроелементи. Аминокиселинни препарати, лекарства за парентерално хранене, когато обичайното е невъзможно.
резюме, добавен на 19.08.2013
Ролята на аминокиселините за човешкото тяло и наследствени нарушения на техния метаболизъм. Фенилкетонурия и форми на заболяването. Честотата на хомоцистинурия и комплексът от нейните характеристики. Хистидинемия: клинични прояви и форми. Биохимична диагностика на левкодистрофия.
резюме, добавен на 11.05.2009
Специално място на протеиновия метаболизъм в разнообразните трансформации на веществата във всички живи организми. Нарушения на биосинтеза и разграждане на протеини в органи и тъкани. Наследствени дефекти в биосинтеза на протеини. Нарушения в екскрецията и крайните етапи на метаболизма на аминокиселините.
резюме, добавен на 22.01.2010г
Описание на фенилкетонурия - наследствено метаболитно заболяване на една от важните аминокиселини (фенилаланин), поради липса или пълно отсъствие на ензим, необходим за метаболизма. Етиология и патогенеза на заболяването, неврологични симптоми, лечение.
презентация, добавена на 15.05.2015
Ролята на черния дроб и бъбреците в протеиновия метаболизъм. Норми на протеини в храненето. Участие на аминокиселините в процесите на биосинтеза и катаболизъм. Тъканна обмяна на нуклеотиди. Синтез и катаболизъм на ДНК и РНК. Регулиране на процесите на азотен метаболизъм. Патология на азотния метаболизъм.
курсова работа, добавена на 12/06/2008
Запознаване с понятието, същността и процесите на обмяната на веществата. Разглеждане на особеностите на създаването на молекули на аминокиселини, въглехидрати, липиди и нуклеинови киселини. Образуването на всички клетки и тъкани, освобождаването на енергия в процеса на обмяната на веществата в тялото.
презентация, добавена на 02.06.2015
Класификация и клинични прояви на метаболитни нарушения. Наследствени метаболитни нарушения. Разпространението на наследствени метаболитни заболявания с неонатален дебют. Клинична характеристика на вродени метаболитни дефекти.
презентация, добавена на 03.07.2015
Ролята на клетъчните органели в енергийните процеси, нервните клетки. Метаболизъм на въглехидратите и особености на енергийното снабдяване на мозъка. Метаболизъм на липиди, протеини и аминокиселини. Ролята на водата за осигуряване на функционирането. Церебрален енергиен метаболизъм.